Anda mungkin juga menyukai
- Application Programmer DNPIDokumen1 halamanApplication Programmer DNPIAlfi RamadhanBelum ada peringkat
- Excel Macro/VBA: Alfi MagfirwanDokumen38 halamanExcel Macro/VBA: Alfi MagfirwanAlfi RamadhanBelum ada peringkat
- Wed Nov 28 13:21:56 2012 Case: E:/My Documents/Kuliah/PGB/LNG Tahap 1a.hsc Flowsheet: Case (Main)Dokumen1 halamanWed Nov 28 13:21:56 2012 Case: E:/My Documents/Kuliah/PGB/LNG Tahap 1a.hsc Flowsheet: Case (Main)Alfi RamadhanBelum ada peringkat
- Curriculum Vitae: Basic InformationDokumen1 halamanCurriculum Vitae: Basic InformationAlfi RamadhanBelum ada peringkat
- Adam Yudhitiyo CivimiDokumen1 halamanAdam Yudhitiyo CivimiAlfi RamadhanBelum ada peringkat
- 1 s2.0 S0024320502021057 MainDokumen7 halaman1 s2.0 S0024320502021057 MainAlfi RamadhanBelum ada peringkat
- Title: Haco Noodle: Hi-Calcium and Protein From Fish Bone WasteDokumen1 halamanTitle: Haco Noodle: Hi-Calcium and Protein From Fish Bone WasteAlfi RamadhanBelum ada peringkat
- Tambahan InventoryDokumen1 halamanTambahan InventoryAlfi RamadhanBelum ada peringkat
- Engineering Progress, Sept. 1967, Pp. 41-47Dokumen1 halamanEngineering Progress, Sept. 1967, Pp. 41-47Alfi RamadhanBelum ada peringkat
- Refference: Pharmaceutical Products. United Kingdom: John Wiley & SonsDokumen1 halamanRefference: Pharmaceutical Products. United Kingdom: John Wiley & SonsAlfi RamadhanBelum ada peringkat
- 8.3.1 Injection Molding MachineDokumen1 halaman8.3.1 Injection Molding MachineAlfi RamadhanBelum ada peringkat
- InventoryDokumen2 halamanInventoryAlfi RamadhanBelum ada peringkat
- OutlineDokumen2 halamanOutlineAlfi RamadhanBelum ada peringkat
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- CH 09Dokumen12 halamanCH 09Daniel DennyBelum ada peringkat
- FSCC 22000 Version 5 and Other RequirementsDokumen29 halamanFSCC 22000 Version 5 and Other Requirementskrishan100% (1)
- Social Stories 1Dokumen23 halamanSocial Stories 1api-472451946Belum ada peringkat
- Contoh Daftar Belanja Obat Praktek PribadiDokumen10 halamanContoh Daftar Belanja Obat Praktek PribadimeryBelum ada peringkat
- Q3-PPT-HEALTH10 - Significance of Global Health InitiativesDokumen20 halamanQ3-PPT-HEALTH10 - Significance of Global Health InitiativesJewel Emerald100% (8)
- Tugas BingDokumen3 halamanTugas BingWahyu YuniarniBelum ada peringkat
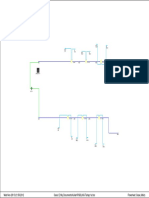
- Transformers Fire Protection: October 2017Dokumen5 halamanTransformers Fire Protection: October 2017Hedi Ben MohamedBelum ada peringkat
- Feet Biomechanics: Mourtzapis Nikolaos Liopetas GeorgiosDokumen16 halamanFeet Biomechanics: Mourtzapis Nikolaos Liopetas GeorgiosgundadanBelum ada peringkat
- GMP History PDFDokumen49 halamanGMP History PDFSubhradipBelum ada peringkat
- Topic 3 Environmental Risk AssessmentDokumen47 halamanTopic 3 Environmental Risk Assessment郭思凱Belum ada peringkat
- Evening Street Review Number 33Dokumen180 halamanEvening Street Review Number 33Barbara Bergmann0% (1)
- The Management of Psychiatric Emergencies: MedicineDokumen9 halamanThe Management of Psychiatric Emergencies: MedicineAnggi CalapiBelum ada peringkat
- Hospital Designing and PlanningDokumen55 halamanHospital Designing and PlanningMukesh DubeyBelum ada peringkat
- Rallygram 2022 FinalDokumen4 halamanRallygram 2022 Finalapi-654829982Belum ada peringkat
- Luyando Nkwemu's Field ProjectDokumen14 halamanLuyando Nkwemu's Field ProjectMathews SikasoteBelum ada peringkat
- Method Development and Validation of RP-HPLCDokumen8 halamanMethod Development and Validation of RP-HPLCManuel PardiñaBelum ada peringkat
- Esophagus Plan Comparison-Supafirefly Vs ImrtDokumen17 halamanEsophagus Plan Comparison-Supafirefly Vs Imrtapi-602263051Belum ada peringkat
- WI-962353 QEL03 Audit Plan LetterDokumen3 halamanWI-962353 QEL03 Audit Plan LetterChakky DestinyBelum ada peringkat
- National Political Parties in India: Hidayatullah National Law University Raipur, ChhattisgarhDokumen34 halamanNational Political Parties in India: Hidayatullah National Law University Raipur, ChhattisgarhshiviBelum ada peringkat
- Manual For Doctors To Evaluate Permanent Physical ImpairmentDokumen33 halamanManual For Doctors To Evaluate Permanent Physical Impairmentyuvrajavi100% (1)
- The Mind in Homeopathy: Rhetoric vs. RealityDokumen6 halamanThe Mind in Homeopathy: Rhetoric vs. RealitySaurav AroraBelum ada peringkat
- ROTC ReviewerDokumen6 halamanROTC ReviewerJenelyn BorbonBelum ada peringkat
- Alimentos PH Urinario GatosDokumen4 halamanAlimentos PH Urinario GatosLarissa VianaBelum ada peringkat
- Unlock Your Tight Hip FlexorsDokumen63 halamanUnlock Your Tight Hip FlexorsPina Olson Campbell100% (2)
- Ammonium ChlorideDokumen3 halamanAmmonium ChlorideJuan David BastidasBelum ada peringkat
- Medical TattooingDokumen66 halamanMedical TattooingTattooTattoo100% (4)
- Aaha Dental Guidelines PDFDokumen21 halamanAaha Dental Guidelines PDFGianfranco DanziBelum ada peringkat
- Vampires and Death in New England, 1784 To 1892Dokumen17 halamanVampires and Death in New England, 1784 To 1892Ferencz IozsefBelum ada peringkat
- OkhlaDokumen55 halamanOkhlaPoojit Popli50% (2)
- 10 Simulation Exercises As A Patient Safety Strategy PDFDokumen9 halaman10 Simulation Exercises As A Patient Safety Strategy PDFAmanda DavisBelum ada peringkat