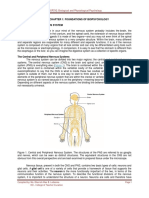
Anda mungkin juga menyukai
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesDari EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesBelum ada peringkat
- Neuron and Glial CellsDokumen8 halamanNeuron and Glial CellsMurilo Bonet100% (1)
- Histology: Neuron Cells Types and StructureDokumen7 halamanHistology: Neuron Cells Types and StructureAli HayderBelum ada peringkat
- Nervous System: The Dog Stole the Professor's NotesDari EverandNervous System: The Dog Stole the Professor's NotesBelum ada peringkat
- Nerve Tissue & the Nervous System IntroductionDokumen21 halamanNerve Tissue & the Nervous System IntroductionDan UvarovBelum ada peringkat
- Mind at Rest: How Neuron Structure Evolves in the Sleep Cycle.Dari EverandMind at Rest: How Neuron Structure Evolves in the Sleep Cycle.Belum ada peringkat
- N 4Dokumen85 halamanN 4Riya ThakkarBelum ada peringkat
- 09 Nerve TissueDokumen13 halaman09 Nerve TissueGovind V PBelum ada peringkat
- Assessment 8Dokumen3 halamanAssessment 8Dianne LarozaBelum ada peringkat
- Nervous Tissue: Prof. Dr. Anas Sarwar QureshiDokumen12 halamanNervous Tissue: Prof. Dr. Anas Sarwar QureshiShafaqat Ghani Shafaqat GhaniBelum ada peringkat
- Nervous System: Communications NetworkDokumen22 halamanNervous System: Communications NetworkAaron TilAhunBelum ada peringkat
- Nervous System ResonanceDokumen72 halamanNervous System ResonanceEkta ManglaniBelum ada peringkat
- Nervous System Structure and FunctionDokumen12 halamanNervous System Structure and FunctionLeon MarkoBelum ada peringkat
- Lecture 9 Histology College of Education34351Dokumen6 halamanLecture 9 Histology College of Education34351Zelal Doski100% (1)
- Nervous Tissue Structure and FunctionDokumen4 halamanNervous Tissue Structure and FunctionFadhil Hussam AhmedBelum ada peringkat
- Animal Nervous System: Logo HereDokumen32 halamanAnimal Nervous System: Logo HereAnne Claudette Capin TeofiloBelum ada peringkat
- Nerve & MuscleDokumen82 halamanNerve & MusclebookaccountBelum ada peringkat
- Nervous SystemDokumen23 halamanNervous Systemde_me3Belum ada peringkat
- Week 5Dokumen26 halamanWeek 5Avril Ros ReyesBelum ada peringkat
- The Nervous System 2Dokumen84 halamanThe Nervous System 2Erika MatiasBelum ada peringkat
- Acknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsDokumen7 halamanAcknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsAndreea LăzăroiuBelum ada peringkat
- Untitled DocumentDokumen15 halamanUntitled DocumentmaBelum ada peringkat
- Nervous TissueDokumen6 halamanNervous TissueAhmed JawdetBelum ada peringkat
- The Nervous System Is Divided Into Two PartsDokumen5 halamanThe Nervous System Is Divided Into Two PartsTahir AzizBelum ada peringkat
- neuroconductivityDokumen16 halamanneuroconductivitysandrajoshy06Belum ada peringkat
- Nervous System - Histology - CompressedDokumen15 halamanNervous System - Histology - Compressedadlin munirahBelum ada peringkat
- Psy 413 ProjectDokumen38 halamanPsy 413 ProjectDumebi AneneBelum ada peringkat
- 1 Hist Sept 13 ReadingNerve - Tissue2020Dokumen26 halaman1 Hist Sept 13 ReadingNerve - Tissue2020n-gorBelum ada peringkat
- Understanding Nervous TissueDokumen30 halamanUnderstanding Nervous TissueOtencianoBelum ada peringkat
- Nervous System HistologyDokumen4 halamanNervous System HistologyShelly Stephanie Bintoro100% (1)
- Anatomy of Nervous TissueDokumen54 halamanAnatomy of Nervous TissueNand PrakashBelum ada peringkat
- Macroglia+Neuron ASG 16-01-2023 PDFDokumen9 halamanMacroglia+Neuron ASG 16-01-2023 PDFRajat AgrawalBelum ada peringkat
- The Nervous SystemDokumen36 halamanThe Nervous SystemAebie AquinoBelum ada peringkat
- Samenvatting Hoofdstuk 3 B&CDokumen3 halamanSamenvatting Hoofdstuk 3 B&CMarlijn Ter MorsBelum ada peringkat
- Nervous TissueDokumen25 halamanNervous TissueSumayya KabeerBelum ada peringkat
- Veterinary Neurobiology: Class NotesDokumen101 halamanVeterinary Neurobiology: Class NotesMaximilian AlexandruBelum ada peringkat
- Nerve Muscle PhysioDokumen30 halamanNerve Muscle Physiopatel_hanisha06Belum ada peringkat
- Nerve-Part 1Dokumen30 halamanNerve-Part 1GiftBelum ada peringkat
- Navigation Search Human Brain Brain (Disambiguation)Dokumen11 halamanNavigation Search Human Brain Brain (Disambiguation)keola greavesBelum ada peringkat
- Understanding the nervous systemDokumen52 halamanUnderstanding the nervous systemKathy Angeli Bel-idaBelum ada peringkat
- Itzar - Neurohistology PDFDokumen186 halamanItzar - Neurohistology PDFahmad aqilBelum ada peringkat
- Anfis System NeurologiDokumen51 halamanAnfis System Neurologianon_822636748Belum ada peringkat
- The Nervous TissueDokumen11 halamanThe Nervous TissuelikeleliletabaBelum ada peringkat
- Three Main Functions:: The Nervous System Is The Master Controlling System of The BodyDokumen34 halamanThree Main Functions:: The Nervous System Is The Master Controlling System of The Bodyapi-19641337Belum ada peringkat
- The Nervous SystemDokumen5 halamanThe Nervous SystemStephen YorBelum ada peringkat
- Anatomy and PhysiologyDokumen18 halamanAnatomy and PhysiologyMaggyBelum ada peringkat
- Table of Content: Serial No Page NoDokumen33 halamanTable of Content: Serial No Page NoSID RAJPOOTBelum ada peringkat
- Bio and PhysioDokumen42 halamanBio and PhysioApenton MimiBelum ada peringkat
- Basic AnotomyDokumen48 halamanBasic AnotomyTadiwa MusaraBelum ada peringkat
- Nerve TissueDokumen5 halamanNerve TissueSuccessfactors TrainerBelum ada peringkat
- Neuroanatomy: Morphology of The BrainDokumen19 halamanNeuroanatomy: Morphology of The BrainAsad_Siddiqi_1076Belum ada peringkat
- University of Pennsylvania Neuroscience ResearchDokumen25 halamanUniversity of Pennsylvania Neuroscience Researchapi-495461280Belum ada peringkat
- NERVOUS TISSUE STRUCTURE AND FUNCTIONDokumen28 halamanNERVOUS TISSUE STRUCTURE AND FUNCTIONIbrahim BabatundeBelum ada peringkat
- Second Quarter Grade 10 - Science HANDOUT No.2 On Neurons, Reflex, Reflex Arc & Brain General Information About The Nervous SystemDokumen2 halamanSecond Quarter Grade 10 - Science HANDOUT No.2 On Neurons, Reflex, Reflex Arc & Brain General Information About The Nervous SystemMartin ReaporBelum ada peringkat
- Unit 5 The Nervous System: Learning OutcomesDokumen11 halamanUnit 5 The Nervous System: Learning OutcomesDeolita BadiangBelum ada peringkat
- Nervous TissueDokumen101 halamanNervous TissueMitzel SapaloBelum ada peringkat
- Summary NeurobiologyDokumen42 halamanSummary NeurobiologytjBelum ada peringkat
- Nervous Tissue Functions and StructuresDokumen3 halamanNervous Tissue Functions and StructuresAlbertBelum ada peringkat
- 9SUMMARYNERVOUSSYSTEMDokumen24 halaman9SUMMARYNERVOUSSYSTEMArvenBitasBelum ada peringkat
- El Conectome Humano - Olaf Sporns (ING)Dokumen21 halamanEl Conectome Humano - Olaf Sporns (ING)Toñitoo H. SkelterBelum ada peringkat
- Hypersensitivity ReactionDokumen36 halamanHypersensitivity ReactionMohana Sundaram100% (1)
- Practical Applications: Crown Lengthening RevisitedDokumen7 halamanPractical Applications: Crown Lengthening RevisitedKyoko CPBelum ada peringkat
- Incomplete ossification of carpal, tarsal and navicular bones in dysmature foalDokumen7 halamanIncomplete ossification of carpal, tarsal and navicular bones in dysmature foalNathalia PedrazaBelum ada peringkat
- Early mobilization effect on postpartum uterine involutionDokumen5 halamanEarly mobilization effect on postpartum uterine involutionElis FatmayantiBelum ada peringkat
- Ncert Solutions Class 8 Science Chapter 10Dokumen10 halamanNcert Solutions Class 8 Science Chapter 10JollyBelum ada peringkat
- Grade 8 Digestive System LessonDokumen4 halamanGrade 8 Digestive System Lessonchristian jade quijano100% (4)
- Normal OcclusionDokumen33 halamanNormal OcclusionFatimah Az-ZahrahBelum ada peringkat
- Massive Transfusion and Massive Transfusion ProtocolDokumen9 halamanMassive Transfusion and Massive Transfusion ProtocolRamachandran SundararamanBelum ada peringkat
- ABO Grouping BB LabDokumen5 halamanABO Grouping BB LabBless MarieBelum ada peringkat
- A Systematic Review of Oral Myofunctional TherapyDokumen12 halamanA Systematic Review of Oral Myofunctional TherapyDiana CaperaBelum ada peringkat
- Division of Spinal CordDokumen23 halamanDivision of Spinal CordBibek SutradharBelum ada peringkat
- Vital Signs and Physical Exam FindingsDokumen30 halamanVital Signs and Physical Exam Findingssamangelo94Belum ada peringkat
- Atherosclerosis: A Brief SummaryDokumen7 halamanAtherosclerosis: A Brief Summarynan doeBelum ada peringkat
- Life Processes 2Dokumen30 halamanLife Processes 2Generic nameBelum ada peringkat
- Acute GastritisDokumen43 halamanAcute GastritisANCHAL SHARMABelum ada peringkat
- Anti Tetanus SerumDokumen3 halamanAnti Tetanus SerumNoah Kent MojicaBelum ada peringkat
- Overview of The Endocrine System: Lesson A2.1Dokumen35 halamanOverview of The Endocrine System: Lesson A2.1Khobie PabilicoBelum ada peringkat
- Catch Up Friday Feb 29Dokumen4 halamanCatch Up Friday Feb 29leicatapangBelum ada peringkat
- Anatomy and Physiology-A ReviewDokumen38 halamanAnatomy and Physiology-A Reviewjava_biscocho1229100% (4)
- KS3 Human Organ SystemDokumen2 halamanKS3 Human Organ Systemejljn1Belum ada peringkat
- Circulatory system microanatomyDokumen5 halamanCirculatory system microanatomyJohnny BravoBelum ada peringkat
- Nakamura IIDokumen10 halamanNakamura IIMairaMaraviChavezBelum ada peringkat
- External Feature of The Insect: Prepared By: Ghada Abou EleineinDokumen32 halamanExternal Feature of The Insect: Prepared By: Ghada Abou EleineinAlham MohammadBelum ada peringkat
- LKPD Digestive-System PDFDokumen3 halamanLKPD Digestive-System PDFLala SiibolaBelum ada peringkat
- ORMCO Case StudiesDokumen36 halamanORMCO Case StudiesDominikaSkórkaBelum ada peringkat
- Ahmad KamsyaDokumen9 halamanAhmad Kamsyaraynhard b. fandresBelum ada peringkat
- NVBSP BMR Forms (BM 01-10) - Revised2022Dokumen23 halamanNVBSP BMR Forms (BM 01-10) - Revised2022lynore sandovalBelum ada peringkat
- Kidney, Suprarenal Gland, Ureter and Bladder AnatomyDokumen46 halamanKidney, Suprarenal Gland, Ureter and Bladder AnatomyzahrazhfBelum ada peringkat
- Blue BrainDokumen3 halamanBlue Braingiulio141091Belum ada peringkat
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDari Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessPenilaian: 4 dari 5 bintang4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDari EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RacePenilaian: 4.5 dari 5 bintang4.5/5 (515)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDari EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeBelum ada peringkat
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDari EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindPenilaian: 4.5 dari 5 bintang4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDari EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondPenilaian: 4 dari 5 bintang4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyDari EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyPenilaian: 3.5 dari 5 bintang3.5/5 (31)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDari EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionPenilaian: 4 dari 5 bintang4/5 (811)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDari EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesPenilaian: 4.5 dari 5 bintang4.5/5 (396)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDari EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorBelum ada peringkat
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDari EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldPenilaian: 4.5 dari 5 bintang4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDari EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedPenilaian: 4 dari 5 bintang4/5 (11)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildDari EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildPenilaian: 4.5 dari 5 bintang4.5/5 (44)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineDari EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestinePenilaian: 4 dari 5 bintang4/5 (17)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishDari EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishPenilaian: 4 dari 5 bintang4/5 (30)
- Summary of Robert M. Sapolsky's DeterminedDari EverandSummary of Robert M. Sapolsky's DeterminedPenilaian: 5 dari 5 bintang5/5 (2)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDari EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsPenilaian: 4.5 dari 5 bintang4.5/5 (4)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesDari EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesPenilaian: 3.5 dari 5 bintang3.5/5 (56)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceDari EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceBelum ada peringkat
- Gathering Moss: A Natural and Cultural History of MossesDari EverandGathering Moss: A Natural and Cultural History of MossesPenilaian: 4.5 dari 5 bintang4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsDari EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsBelum ada peringkat
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationDari EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationPenilaian: 4 dari 5 bintang4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeDari EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifePenilaian: 4 dari 5 bintang4/5 (523)
- Superlative: The Biology of ExtremesDari EverandSuperlative: The Biology of ExtremesPenilaian: 4.5 dari 5 bintang4.5/5 (51)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDari EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsPenilaian: 4.5 dari 5 bintang4.5/5 (2083)
- Lymph & Longevity: The Untapped Secret to HealthDari EverandLymph & Longevity: The Untapped Secret to HealthPenilaian: 4.5 dari 5 bintang4.5/5 (13)