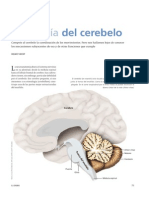
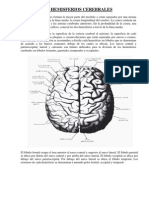
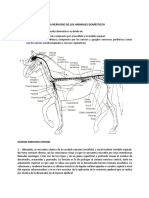
Anda mungkin juga menyukai
- Corteza CerebelosaDokumen11 halamanCorteza CerebelosaLucía FaríasBelum ada peringkat
- Barrera HematoencefalicaDokumen6 halamanBarrera HematoencefalicaVanessa Frisancho0% (1)
- Dialnet SubstanciaReticular 6293548Dokumen30 halamanDialnet SubstanciaReticular 6293548ALIDA MARINA AGUILAR GUTIERREZBelum ada peringkat
- NeurocienciasDokumen17 halamanNeurocienciasYoselin Valentina CastañoBelum ada peringkat
- Cerebelo y Plexos CoroideosDokumen59 halamanCerebelo y Plexos CoroideosJackie RoblesBelum ada peringkat
- Ce RebeloDokumen14 halamanCe RebeloMaferAlonsoBelum ada peringkat
- Nervio FacialDokumen18 halamanNervio FacialAguus CabreraBelum ada peringkat
- Anatomía e Histología de La RetinaDokumen10 halamanAnatomía e Histología de La RetinaReiniel Díaz Pérez100% (1)
- Bulbo RaquídeoDokumen7 halamanBulbo RaquídeoJoha VelasquezBelum ada peringkat
- Capítulo39 T2 2 ParteDokumen87 halamanCapítulo39 T2 2 ParteYasmira CerrónBelum ada peringkat
- MesencefaloDokumen17 halamanMesencefaloClinton anderson Luque mamaniBelum ada peringkat
- Histologia - TEJIDO NERVIOSODokumen4 halamanHistologia - TEJIDO NERVIOSOFernando Huamani HuarotoBelum ada peringkat
- Seram2014 S-0609Dokumen35 halamanSeram2014 S-0609DANIELABelum ada peringkat
- Apunte Cerebelo 2010Dokumen14 halamanApunte Cerebelo 2010Marlyns Jovanna Valecillos CastilloBelum ada peringkat
- Tejido Nervioso y Sistema NerviosoDokumen16 halamanTejido Nervioso y Sistema Nerviosomsolorzano-1Belum ada peringkat
- Disección cerebro resDokumen9 halamanDisección cerebro resLizette AngelBelum ada peringkat
- Núcleo celular y cromatinaDokumen18 halamanNúcleo celular y cromatinaJesus Montemayor BeltranBelum ada peringkat
- 1 MAT COMPL de SNC y CerebeloDokumen4 halaman1 MAT COMPL de SNC y CerebeloNelly DiazBelum ada peringkat
- Función y anatomía del cerebeloDokumen15 halamanFunción y anatomía del cerebeloursula142100% (1)
- Brodman AreasDokumen36 halamanBrodman AreasabiBelum ada peringkat
- Anatomía Del CerebeloDokumen3 halamanAnatomía Del CerebeloMila CcasaniBelum ada peringkat
- Anatomia Del Desarrollo Del Sistema Nervioso - Abril PreatoniDokumen20 halamanAnatomia Del Desarrollo Del Sistema Nervioso - Abril PreatoniGloria Solis San Martin100% (1)
- Resume NDokumen24 halamanResume NDINA REBECA FUENTES PÉREZBelum ada peringkat
- Anatomía Cerebro HistologíaDokumen6 halamanAnatomía Cerebro HistologíaMarlenMtz100% (1)
- Histología Del CerebeloDokumen32 halamanHistología Del CerebeloanaBelum ada peringkat
- El Cerebelo (Modificado x1)Dokumen7 halamanEl Cerebelo (Modificado x1)albarranleivarossydarianaBelum ada peringkat
- Investigacion Del TimoDokumen4 halamanInvestigacion Del TimoYairely Perez GallegosBelum ada peringkat
- Sistema Nervioso ResumenDokumen6 halamanSistema Nervioso ResumenSofía ColzeraBelum ada peringkat
- Hemisferios Cerebrales - Corteza Cerebral Celulas Capas Localizacion Funcional de La...Dokumen8 halamanHemisferios Cerebrales - Corteza Cerebral Celulas Capas Localizacion Funcional de La...Annyta CervantesBelum ada peringkat
- Cápsula InternaDokumen5 halamanCápsula InternaAnthony Romero100% (2)
- Cerebelo. Neuro-2Dokumen15 halamanCerebelo. Neuro-2Gabriel DavalosBelum ada peringkat
- Separata Sistema NerviosoDokumen17 halamanSeparata Sistema NerviosoVíctor Andres Mateo SanchezBelum ada peringkat
- Atlas Histología Vegetal y AnimalDokumen27 halamanAtlas Histología Vegetal y AnimalQuien S SabeBelum ada peringkat
- Cerebelo Microanatomia IDokumen29 halamanCerebelo Microanatomia IAle MedranoBelum ada peringkat
- Embiologia: Sistema NerviosoDokumen96 halamanEmbiologia: Sistema NerviosoBelen Muñoz RománBelum ada peringkat
- Sistema nervioso: análisis histológico de la corteza cerebral, tálamo e hipocampo en caso clínicoDokumen7 halamanSistema nervioso: análisis histológico de la corteza cerebral, tálamo e hipocampo en caso clínicoJuan RodriguesBelum ada peringkat
- Capítulo de Atlas Citología e Histología 2005 Wolfgang PDFDokumen12 halamanCapítulo de Atlas Citología e Histología 2005 Wolfgang PDFHannie BertalotBelum ada peringkat
- Cerebelo: Estructura y Función del Órgano del Equilibrio y Coordinación MotoraDokumen8 halamanCerebelo: Estructura y Función del Órgano del Equilibrio y Coordinación MotoraAlfonsina AcostaBelum ada peringkat
- Núcleo celular: estructura y funciónDokumen6 halamanNúcleo celular: estructura y funciónAnonymous VvMA4CTDBelum ada peringkat
- HistologiÌ A I Segundo ParcialDokumen13 halamanHistologiÌ A I Segundo Parcial2303542Belum ada peringkat
- Atlas de Histologà A Descriptiva 2012 PDFDokumen5 halamanAtlas de Histologà A Descriptiva 2012 PDFakuaBelum ada peringkat
- Histología de la corteza cerebralDokumen8 halamanHistología de la corteza cerebralRaymundo PargaBelum ada peringkat
- Desarrollo Del Sistema NerviosoDokumen18 halamanDesarrollo Del Sistema NerviosoKaterine AvilaBelum ada peringkat
- CrustáceosDokumen53 halamanCrustáceosFarithstiverson Cristancho SierraBelum ada peringkat
- Corteza CerebralDokumen6 halamanCorteza CerebralEmily PerezBelum ada peringkat
- Nucleo Celular - BiologiaDokumen17 halamanNucleo Celular - BiologiaDeisy Magdalena Morinigo UlderaBelum ada peringkat
- Tejidos sanguíneo y nervioso: clasificación, estructura y funciónDokumen150 halamanTejidos sanguíneo y nervioso: clasificación, estructura y funciónLuis Enrique Vera CamposBelum ada peringkat
- Embriologia Medica Guia de Practicas Cap 3 CarlosDokumen33 halamanEmbriologia Medica Guia de Practicas Cap 3 CarlosLaura Orellana100% (2)
- Nucleo CelularDokumen14 halamanNucleo Celularl1l2Belum ada peringkat
- Nucleo CelularDokumen33 halamanNucleo Celularsirhecrulle100% (5)
- Instituto Universitario de Ciencias de La Salud Fundacion Héctor A. Barceló Facultad de MedicinaDokumen6 halamanInstituto Universitario de Ciencias de La Salud Fundacion Héctor A. Barceló Facultad de MedicinaSamuel RomeroBelum ada peringkat
- Tejido Nervioso Cuestionario - Sheila MendozaDokumen7 halamanTejido Nervioso Cuestionario - Sheila MendozaShei MendozaBelum ada peringkat
- CEFALOCORDADOSDokumen20 halamanCEFALOCORDADOSJhon Contreras75% (4)
- 4. Configuración interna de tronco encefalicoDokumen23 halaman4. Configuración interna de tronco encefalicoWilliam HernándezBelum ada peringkat
- Neuroanatomia Capitulo 6 Al 10Dokumen166 halamanNeuroanatomia Capitulo 6 Al 10lizzyrodnuzBelum ada peringkat
- Resumen de Cerebelo.Dokumen8 halamanResumen de Cerebelo.Daniel Velásquez CamposBelum ada peringkat
- Taller AnatomíaDokumen13 halamanTaller AnatomíaCarolinaBelum ada peringkat
- Neurofisiologia Del EncefaloDokumen31 halamanNeurofisiologia Del EncefaloJulian DavidBelum ada peringkat
- Análisis Del Riesgo en La Evaluación de ProyectosDokumen17 halamanAnálisis Del Riesgo en La Evaluación de Proyectospatty41Belum ada peringkat
- Epiescleritis: Ojo rojo, afectación limitada a área temporal con aspecto rojo brillanteDokumen6 halamanEpiescleritis: Ojo rojo, afectación limitada a área temporal con aspecto rojo brillantepatty41100% (1)
- Anatomia RandomDokumen45 halamanAnatomia Randompatty41Belum ada peringkat
- GlaucomaDokumen7 halamanGlaucomapatty41Belum ada peringkat
- Medio ambiente laboral, factores de riesgo y prevenciónDokumen22 halamanMedio ambiente laboral, factores de riesgo y prevenciónRosa Elena Crisanto VelascoBelum ada peringkat
- Urología Villamedic 2016 - Diapositivas de VideoclaseDokumen95 halamanUrología Villamedic 2016 - Diapositivas de VideoclaseFranz Arturo Lozada Hernández100% (1)
- Salud y SociedadDokumen15 halamanSalud y Sociedadpatty41Belum ada peringkat
- Eliminacion Aguas ResidualesDokumen19 halamanEliminacion Aguas Residualespatty41Belum ada peringkat
- Sindrome de Hipertension IntracranealDokumen59 halamanSindrome de Hipertension Intracranealbrandoc93Belum ada peringkat
- Análisis Del Riesgo en La Evaluación de ProyectosDokumen17 halamanAnálisis Del Riesgo en La Evaluación de Proyectospatty41Belum ada peringkat
- Edema de ReinkeDokumen18 halamanEdema de Reinkepatty41Belum ada peringkat
- Análisis de Proyectos Con Distinta Vida Útil PDFDokumen7 halamanAnálisis de Proyectos Con Distinta Vida Útil PDFpatty41Belum ada peringkat
- Análisis Del Riesgo en Proyectos 2016 IIDokumen9 halamanAnálisis Del Riesgo en Proyectos 2016 IIpatty41Belum ada peringkat
- Soap 2Dokumen1 halamanSoap 2patty41Belum ada peringkat
- Pruebas Diagnosticas2Dokumen6 halamanPruebas Diagnosticas2p1e2d3Belum ada peringkat
- Antibioticoterapia y Analgesicos en Cirugia 2019Dokumen29 halamanAntibioticoterapia y Analgesicos en Cirugia 2019patty41Belum ada peringkat
- Salud Y Sociedad Iii S018 - I: Sesión 4: Razones - Proporciones - TasasDokumen15 halamanSalud Y Sociedad Iii S018 - I: Sesión 4: Razones - Proporciones - Tasaspatty41Belum ada peringkat
- Histología Semana 5Dokumen41 halamanHistología Semana 5patty41Belum ada peringkat
- Asma Bronquial FarmacologíaDokumen36 halamanAsma Bronquial Farmacologíapatty41Belum ada peringkat
- Lectura OptativaDokumen2 halamanLectura Optativapatty41Belum ada peringkat
- Guía Práctica de Manejo Clínico Europea de Otitis Media Aguda PDFDokumen16 halamanGuía Práctica de Manejo Clínico Europea de Otitis Media Aguda PDFpatty41Belum ada peringkat
- AR Guía EuropeaDokumen16 halamanAR Guía Europeapatty41Belum ada peringkat
- Ciclo CelularDokumen15 halamanCiclo Celularpatty41Belum ada peringkat
- Inmunología Del EmbarazoDokumen20 halamanInmunología Del Embarazopatty41Belum ada peringkat
- Perú en Números 2014Dokumen200 halamanPerú en Números 2014Gianfrancoqf0% (1)
- Cartilla 2.PDF ProteccionDokumen2 halamanCartilla 2.PDF Proteccionpatty41Belum ada peringkat
- Tejido ConectivoDokumen129 halamanTejido Conectivopatty41Belum ada peringkat
- Sistema de ComplementoDokumen45 halamanSistema de Complementopatty41Belum ada peringkat
- Trabajo de PartoDokumen72 halamanTrabajo de Partopatty41Belum ada peringkat
- Guia # 1 Grado 7 Segundo PeriodoDokumen8 halamanGuia # 1 Grado 7 Segundo PeriodoMilena CamachoBelum ada peringkat
- Libreto Amor Sin Barreras CortaDokumen49 halamanLibreto Amor Sin Barreras CortaLuis Layc60% (5)
- Configuración de plotters y estilos de trazado en AutoCAD 2000Dokumen12 halamanConfiguración de plotters y estilos de trazado en AutoCAD 2000Oscar PichardoBelum ada peringkat
- Prueba - Amigos en El Bosque - Formularios de GoogleDokumen9 halamanPrueba - Amigos en El Bosque - Formularios de GoogleNataliaMackarenaVeraFuentesBelum ada peringkat
- Sesión 05 - Tilde Diacrítica-Juego de MemoriaDokumen25 halamanSesión 05 - Tilde Diacrítica-Juego de MemoriaMasterDefcomBelum ada peringkat
- Pia AdministraciónDokumen5 halamanPia AdministraciónPerla Andrea Mendoza RodríguezBelum ada peringkat
- 1BTOF Royo Claudia Auditoria Digital PDFDokumen2 halaman1BTOF Royo Claudia Auditoria Digital PDFCLAUDIA ROYOBelum ada peringkat
- Un Paso Hacia La Paz ColombiaDokumen1 halamanUn Paso Hacia La Paz ColombiaPaola SerranoBelum ada peringkat
- Fotografia Jaen Siglo XixDokumen3 halamanFotografia Jaen Siglo XixRosa XiaoBelum ada peringkat
- Hotel Best Western Plus Santa Marta 2023Dokumen1 halamanHotel Best Western Plus Santa Marta 2023Sergio SantanillaBelum ada peringkat
- Ante Tu AltarDokumen1 halamanAnte Tu AltarReynel AlexandroBelum ada peringkat
- Wiitdb EsDokumen16 halamanWiitdb Esluis cBelum ada peringkat
- Lecturas Progresivas Vocales y Minúsculas 2019Dokumen31 halamanLecturas Progresivas Vocales y Minúsculas 2019Daniela Cuevas RomoBelum ada peringkat
- EXAMEN DE LITERATURA PERUANADokumen3 halamanEXAMEN DE LITERATURA PERUANAroleiverBelum ada peringkat
- Ensayo Con Analisis de La Pelicula El Tigre y El DragonDokumen6 halamanEnsayo Con Analisis de La Pelicula El Tigre y El DragonYovani ChulloBelum ada peringkat
- Domo TicaDokumen3 halamanDomo TicaDavid Poma Q.Belum ada peringkat
- ANALISIS CRÍTICO El Caballero de La Armadura OxidadaDokumen4 halamanANALISIS CRÍTICO El Caballero de La Armadura OxidadaMartaBelum ada peringkat
- Cuento NarrativoDokumen2 halamanCuento Narrativorocio chinkimBelum ada peringkat
- Planilla de Andamio WordDokumen1 halamanPlanilla de Andamio WordMarquitosGil0% (1)
- Simón y El Carro de FuegoDokumen5 halamanSimón y El Carro de Fuegosebas2060% (1)
- Ejercicio 1Dokumen5 halamanEjercicio 1Daniela Tkm0% (2)
- Diez Reglas para Deslumbrar en Una Entrevista de TrabajoDokumen2 halamanDiez Reglas para Deslumbrar en Una Entrevista de TrabajoSilvia6tmaBelum ada peringkat
- Las FábulasDokumen2 halamanLas FábulasMaria Luz CortegosoBelum ada peringkat
- PLAN DE CLASE N°3 Fiesta de Los EstudiantesDokumen3 halamanPLAN DE CLASE N°3 Fiesta de Los EstudiantesFernando garecaBelum ada peringkat
- Hoja de VidaDokumen5 halamanHoja de VidaMaria Lucia ViciniBelum ada peringkat
- Mito de PerséfoneDokumen7 halamanMito de PerséfoneGeorge AdivinaBelum ada peringkat
- Armado Del Motor D6DDokumen43 halamanArmado Del Motor D6DJose Garcia Rodriguez86% (7)
- Consume - Maverick City Musica Ft. Laila Olvera 6 Nate Díaz)Dokumen5 halamanConsume - Maverick City Musica Ft. Laila Olvera 6 Nate Díaz)Mónica ChávezBelum ada peringkat
- El Lapiz MagicoDokumen2 halamanEl Lapiz MagicoKaren Margarita100% (1)
- Análisis de personajes en obras dramáticasDokumen3 halamanAnálisis de personajes en obras dramáticasFernando DíazBelum ada peringkat