Anda mungkin juga menyukai
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Graf DistributionDokumen3 halamanGraf DistributionLyna NaderBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Postoperative CareDokumen41 halamanPostoperative CareLyna NaderBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Student Binding ContractDokumen3 halamanStudent Binding ContractLyna NaderBelum ada peringkat
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- f2 MatematikDokumen1 halamanf2 MatematikLyna NaderBelum ada peringkat
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Igcse 0606Dokumen1 halamanIgcse 0606Lyna NaderBelum ada peringkat
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Revision Module Midterm Maths Form 5Dokumen19 halamanRevision Module Midterm Maths Form 5Lyna NaderBelum ada peringkat
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Sunset by The BeachDokumen1 halamanSunset by The BeachLyna NaderBelum ada peringkat
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Theory Paper (Bio) SPMDokumen2 halamanTheory Paper (Bio) SPMLyna NaderBelum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Part IIIDokumen7 halamanPart IIILyna NaderBelum ada peringkat
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Atom Proton Number Nucleon NumberDokumen2 halamanAtom Proton Number Nucleon NumberLyna NaderBelum ada peringkat
- Physic Exercise Part 1Dokumen2 halamanPhysic Exercise Part 1Lyna NaderBelum ada peringkat
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Graf DistributionDokumen3 halamanGraf DistributionLyna NaderBelum ada peringkat
- Poem StartDokumen1 halamanPoem StartLyna NaderBelum ada peringkat
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
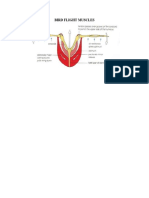
- Bird Flight MusclesDokumen3 halamanBird Flight MusclesLyna NaderBelum ada peringkat
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- Math Pt3 LatihanDokumen2 halamanMath Pt3 LatihanLyna NaderBelum ada peringkat
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Bird Flight MusclesDokumen3 halamanBird Flight MusclesLyna NaderBelum ada peringkat
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Atom Proton Number Nucleon NumberDokumen2 halamanAtom Proton Number Nucleon NumberLyna NaderBelum ada peringkat
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Hello WorldDokumen1 halamanHello WorldLyna NaderBelum ada peringkat
- Add Math Part 1Dokumen4 halamanAdd Math Part 1Lyna NaderBelum ada peringkat
- Hello WorldDokumen1 halamanHello WorldLyna NaderBelum ada peringkat
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Hello WorldDokumen1 halamanHello WorldLyna NaderBelum ada peringkat
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- New Housing Unit Per Unit RM 150 000Dokumen4 halamanNew Housing Unit Per Unit RM 150 000Lyna NaderBelum ada peringkat
- Atom Proton Number Nucleon NumberDokumen2 halamanAtom Proton Number Nucleon NumberLyna NaderBelum ada peringkat
- Part IIIDokumen7 halamanPart IIILyna NaderBelum ada peringkat
- Add Math Part 1Dokumen4 halamanAdd Math Part 1Lyna NaderBelum ada peringkat
- Mathematics Transformations Enlargement SubjectiveDokumen3 halamanMathematics Transformations Enlargement SubjectiveLyna NaderBelum ada peringkat
- Sains Form 2 BM VersionDokumen12 halamanSains Form 2 BM VersionLyna NaderBelum ada peringkat
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Graf DistributionDokumen3 halamanGraf DistributionLyna NaderBelum ada peringkat
- Mathematics Transformations Enlargement SubjectiveDokumen3 halamanMathematics Transformations Enlargement SubjectiveLyna NaderBelum ada peringkat
- Mathematics Transformations Enlargement SubjectiveDokumen2 halamanMathematics Transformations Enlargement SubjectiveLyna NaderBelum ada peringkat
- Quantity Surveyor Dissertation IdeasDokumen8 halamanQuantity Surveyor Dissertation IdeasBestCollegePaperWritingServiceCanada100% (1)
- Format and Guidelines - Dissertation by DR K S SukonDokumen27 halamanFormat and Guidelines - Dissertation by DR K S SukonSar VeshBelum ada peringkat
- Sociolinguistics Help Us To Develop A Better Understanding of Society and Language 1Dokumen6 halamanSociolinguistics Help Us To Develop A Better Understanding of Society and Language 1Ikram KhanBelum ada peringkat
- DIASSDokumen2 halamanDIASSallan basugaBelum ada peringkat
- Prashant Shukla Conduit PPT Case StudyDokumen21 halamanPrashant Shukla Conduit PPT Case StudyPrashant ShuklaBelum ada peringkat
- Two Way AnovaDokumen20 halamanTwo Way AnovaJelly Jean PayeBelum ada peringkat
- American Issues - A Primary Source Reader in United States History.Dokumen3 halamanAmerican Issues - A Primary Source Reader in United States History.MichaelJamesChristopherConklin0% (2)
- Group StructureDokumen2 halamanGroup Structurejade brewBelum ada peringkat
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Cronbach's AlphaDokumen4 halamanCronbach's AlphaRobert RodriguezBelum ada peringkat
- Big Data Analysis GuideDokumen11 halamanBig Data Analysis GuideAnonymous DLEF3GvBelum ada peringkat
- Breadth RequirementsDokumen2 halamanBreadth RequirementsJulian ChoBelum ada peringkat
- Unit 7 Managing Conference and Events Assgt March 2020 PDFDokumen14 halamanUnit 7 Managing Conference and Events Assgt March 2020 PDFDyya EllenaBelum ada peringkat
- The Principle of FunctorialityDokumen15 halamanThe Principle of Functorialitytoschmid007Belum ada peringkat
- Econ 321 Introduction To Statistics Baybars KaracaovaliDokumen3 halamanEcon 321 Introduction To Statistics Baybars Karacaovalilana del reyBelum ada peringkat
- UNIT 4 OBA AssessmentDokumen55 halamanUNIT 4 OBA AssessmentwankamisahBelum ada peringkat
- Psychological Testing: 1.sentence Completion 2. Trending Psychological TestsDokumen17 halamanPsychological Testing: 1.sentence Completion 2. Trending Psychological TestsshehbazBelum ada peringkat
- WSO Global Stroke Factsheet 09-02.2022Dokumen27 halamanWSO Global Stroke Factsheet 09-02.2022Arubayi AbonereBelum ada peringkat
- Design BriefDokumen2 halamanDesign Briefapi-409519942Belum ada peringkat
- Rayons AakashDokumen60 halamanRayons AakashAakash PareekBelum ada peringkat
- Breathing Year 4 ScienceDokumen2 halamanBreathing Year 4 Scienceatiqah zakiBelum ada peringkat
- STS Programme Schedule by Date As at 0407019Dokumen51 halamanSTS Programme Schedule by Date As at 0407019RJ TutorialsBelum ada peringkat
- Utilitarianism: An Ethical Decision Making Approach: February 2019Dokumen15 halamanUtilitarianism: An Ethical Decision Making Approach: February 2019Gary NgBelum ada peringkat
- K073 - SETTVEO - Evidence-Based Reflection and Teacher Development - FINAL - WebDokumen32 halamanK073 - SETTVEO - Evidence-Based Reflection and Teacher Development - FINAL - WebHafezBelum ada peringkat
- Nutritional Assessment of Critically Ill PatientsDokumen8 halamanNutritional Assessment of Critically Ill PatientskikyfauziaBelum ada peringkat
- Classroom Assessment in The K To 12 Basic Education ProgramDokumen50 halamanClassroom Assessment in The K To 12 Basic Education ProgramMaria Luisa Gentoleo QuipitBelum ada peringkat
- Futura Primus Flexible Great Toe FinalDokumen12 halamanFutura Primus Flexible Great Toe FinalNasfikurBelum ada peringkat
- Njala University: Bo Campus-Kowama LocationDokumen32 halamanNjala University: Bo Campus-Kowama LocationALLIEU FB SACCOHBelum ada peringkat
- Applied Research Methods in Social Sciences SyllabusDokumen25 halamanApplied Research Methods in Social Sciences SyllabusNicholas BransonBelum ada peringkat
- Field Study 2Dokumen72 halamanField Study 2Janella DasaBelum ada peringkat
- Program in Zambia: Strengthening Monitoring and Evaluation Systems in The TBDokumen3 halamanProgram in Zambia: Strengthening Monitoring and Evaluation Systems in The TBreem esamBelum ada peringkat
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDari EverandTales from Both Sides of the Brain: A Life in NeurosciencePenilaian: 3 dari 5 bintang3/5 (18)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincDari EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincPenilaian: 3.5 dari 5 bintang3.5/5 (137)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDari EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsPenilaian: 4.5 dari 5 bintang4.5/5 (6)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDari EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RacePenilaian: 4.5 dari 5 bintang4.5/5 (516)
- Gut: the new and revised Sunday Times bestsellerDari EverandGut: the new and revised Sunday Times bestsellerPenilaian: 4 dari 5 bintang4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 4 dari 5 bintang4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Dari EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Penilaian: 4 dari 5 bintang4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDari Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessPenilaian: 4 dari 5 bintang4/5 (33)