Anda mungkin juga menyukai
- Immunointervention in Autoimmune Diseases: Papers Based on an International Meeting in Paris, France, in June 1988Dari EverandImmunointervention in Autoimmune Diseases: Papers Based on an International Meeting in Paris, France, in June 1988J. F. BachBelum ada peringkat
- Fast Facts: Managing Immune-Related Adverse Events in OncologyDari EverandFast Facts: Managing Immune-Related Adverse Events in OncologyBelum ada peringkat
- Interferon Gamma Production in The Course of Mycobacterium Tuberculosis InfectionDokumen9 halamanInterferon Gamma Production in The Course of Mycobacterium Tuberculosis InfectionAndia ReshiBelum ada peringkat
- Abstract PDFDokumen36 halamanAbstract PDFDhini Kartika SariBelum ada peringkat
- Rendahnya Vitd3Dokumen17 halamanRendahnya Vitd3ervina 119260026Belum ada peringkat
- Transplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Dokumen128 halamanTransplant International Special Issue: Abstracts of The 14th Congress of The European Society For Organ Transplantation Volume 22, Issue Supplement s2, Pages 95-222, August 2009Enrique Moreno GonzálezBelum ada peringkat
- Jced 7 E656Dokumen4 halamanJced 7 E656María José VázquezBelum ada peringkat
- Presepsin 6Dokumen189 halamanPresepsin 6donkeyendutBelum ada peringkat
- Research Article: SOCS1 Mimetic Peptide Suppresses Chronic Intraocular Inflammatory Disease (Uveitis)Dokumen16 halamanResearch Article: SOCS1 Mimetic Peptide Suppresses Chronic Intraocular Inflammatory Disease (Uveitis)Paulus SidhartaBelum ada peringkat
- Cytokine: E.S. Robinson, V.P. WerthDokumen9 halamanCytokine: E.S. Robinson, V.P. WerthArthurAkkBelum ada peringkat
- AllergoOncology Opposite Outcomes of Immune Tolerance in Allergy and CancerDokumen13 halamanAllergoOncology Opposite Outcomes of Immune Tolerance in Allergy and CancerallalergexamenBelum ada peringkat
- Secreting CD4 T Cells in Direct Ex Vivo Analysis of Antigen-Specific IFNDokumen10 halamanSecreting CD4 T Cells in Direct Ex Vivo Analysis of Antigen-Specific IFNEdy NoveryBelum ada peringkat
- Human Autoimmune Diseases: A Comprehensive Update: ReviewDokumen27 halamanHuman Autoimmune Diseases: A Comprehensive Update: Reviewkaren rinconBelum ada peringkat
- Research Article: Interleukin-17 Expression in The Barrett's Metaplasia-Dysplasia-Adenocarcinoma SequenceDokumen7 halamanResearch Article: Interleukin-17 Expression in The Barrett's Metaplasia-Dysplasia-Adenocarcinoma SequenceTit fuckerBelum ada peringkat
- Prolactin Increase Frecuency of T Helper FolicularDokumen15 halamanProlactin Increase Frecuency of T Helper Folicularauroragamez2015Belum ada peringkat
- Tosca 1996Dokumen7 halamanTosca 1996HEARTJUGGLERBelum ada peringkat
- Ivermectin For CancerDokumen15 halamanIvermectin For CancerSheena FriesenBelum ada peringkat
- Thesis On Uropathogenic E.coliDokumen7 halamanThesis On Uropathogenic E.colimarialackarlington100% (2)
- Multiple Organ Dysfunction SyndromeDokumen4 halamanMultiple Organ Dysfunction SyndromeAna Reyes100% (1)
- NIH Public Access: Author ManuscriptDokumen8 halamanNIH Public Access: Author ManuscriptJuliet TyeBelum ada peringkat
- Early Expression of Local Cytokines During Systemic Candida Albicans Infection in Murine Intravenous ModelDokumen6 halamanEarly Expression of Local Cytokines During Systemic Candida Albicans Infection in Murine Intravenous Modelcvk717Belum ada peringkat
- 14th ICI Abstract BookDokumen4 halaman14th ICI Abstract BookMuhammadGagasSasongkoBelum ada peringkat
- Candida Albicans: Pathogenicity and Epithelial ImmunityDokumen4 halamanCandida Albicans: Pathogenicity and Epithelial ImmunityIstna Sofia AuliaBelum ada peringkat
- Chochoua COSA Presentation CombinedDokumen8 halamanChochoua COSA Presentation CombinedIma6_Belum ada peringkat
- Out 41Dokumen12 halamanOut 41natanatalisBelum ada peringkat
- Monophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andDokumen14 halamanMonophosphoryl Lipid A Adjuvants in Roles of Aluminum Hydroxide andLeandro CarreñoBelum ada peringkat
- Pulmonary Balantidium Coli Infection in A Leukemic PatientDokumen4 halamanPulmonary Balantidium Coli Infection in A Leukemic PatientNika Dwi AmbarwatiBelum ada peringkat
- Plant Lectins Activate The NLRP3 Inflammasome To Promote Inflammatory DisordersDokumen12 halamanPlant Lectins Activate The NLRP3 Inflammasome To Promote Inflammatory DisordersleonardopinheiroBelum ada peringkat
- Long Live Th2Dokumen18 halamanLong Live Th2ANIS MURNIATIBelum ada peringkat
- ArticuloDokumen9 halamanArticuloShokubutsu AmeBelum ada peringkat
- Cytophagic Histiocytic Panniculitis, Hemophagocytic Lymphohistiocytosis and Undetermined Autoimmune Disorder: Reconciling The PuzzleDokumen5 halamanCytophagic Histiocytic Panniculitis, Hemophagocytic Lymphohistiocytosis and Undetermined Autoimmune Disorder: Reconciling The PuzzleLe DungBelum ada peringkat
- Stress Hyperglycemia and Innate Immune System CellsDokumen9 halamanStress Hyperglycemia and Innate Immune System CellsJuan Fernando BolivarBelum ada peringkat
- Staphy 2Dokumen5 halamanStaphy 2Alexandra MariaBelum ada peringkat
- Review: New Concepts in The Pathophysiology of Inflammatory Bowel DiseaseDokumen11 halamanReview: New Concepts in The Pathophysiology of Inflammatory Bowel DiseaseReynalth Andrew Sinaga100% (1)
- PRIMARY T CELL Keller 2013Dokumen10 halamanPRIMARY T CELL Keller 2013emmanouelaapostolopoulouBelum ada peringkat
- Research Article: SOCS1 Mimetic Peptide Suppresses Chronic Intraocular Inflammatory Disease (Uveitis)Dokumen33 halamanResearch Article: SOCS1 Mimetic Peptide Suppresses Chronic Intraocular Inflammatory Disease (Uveitis)Paulus SidhartaBelum ada peringkat
- Hematopoietic Stem Cell Transplantation Resolves The Immune Deficit Associated With STAT3-Dominant-Negative Hyper-IgE SyndromeDokumen10 halamanHematopoietic Stem Cell Transplantation Resolves The Immune Deficit Associated With STAT3-Dominant-Negative Hyper-IgE SyndromeWalid AissaBelum ada peringkat
- Taller 2. InnataDokumen15 halamanTaller 2. InnataMaría Alejandra Mancera GarcíaBelum ada peringkat
- Bio 1Dokumen3 halamanBio 1anisaBelum ada peringkat
- Clinical and Experimental AllergyDokumen11 halamanClinical and Experimental AllergyrobbyBelum ada peringkat
- Methoxydalbergione Inhibits Esophageal Carcinoma Cell Proliferation and Migration by Inactivating NF‑ΚBDokumen9 halamanMethoxydalbergione Inhibits Esophageal Carcinoma Cell Proliferation and Migration by Inactivating NF‑ΚBRepositorioBelum ada peringkat
- J Infect Dis.-2000-Hang-1738-48Dokumen11 halamanJ Infect Dis.-2000-Hang-1738-48Anonymous hXd6PJlBelum ada peringkat
- Phloretin VivoDokumen9 halamanPhloretin VivoSaud AnsariBelum ada peringkat
- Biomed ..Dokumen10 halamanBiomed ..Romina Molette GonzalesBelum ada peringkat
- Es SIRS o Endotoxemia en El Equino 2014Dokumen15 halamanEs SIRS o Endotoxemia en El Equino 2014Manuela LopezBelum ada peringkat
- Articulo 6 Ip-10okDokumen11 halamanArticulo 6 Ip-10okoscarbio2009Belum ada peringkat
- BMC Clinical Pathology: Protection of Early Phase Hepatic Ischemia-Reperfusion Injury by Cholinergic AgonistsDokumen13 halamanBMC Clinical Pathology: Protection of Early Phase Hepatic Ischemia-Reperfusion Injury by Cholinergic AgonistsPrachi KariaBelum ada peringkat
- Multiple Organ Dysfunction SyndromeDokumen4 halamanMultiple Organ Dysfunction SyndromeSha NaBelum ada peringkat
- Dr. Anang - PKB Bali - 18 Juli 2009 - Short Version - Role of PREBIODokumen45 halamanDr. Anang - PKB Bali - 18 Juli 2009 - Short Version - Role of PREBIOAdhistyRiantoBelum ada peringkat
- Recurrent Mycobacterium Tuberculosis Infection in Systemic Lupus Erythematosus Patient Case Report and ReviewDokumen8 halamanRecurrent Mycobacterium Tuberculosis Infection in Systemic Lupus Erythematosus Patient Case Report and ReviewpaulallenfileBelum ada peringkat
- J Allergy Clin Immunol Volume 135, Number 2Dokumen1 halamanJ Allergy Clin Immunol Volume 135, Number 2Tommy LiuBelum ada peringkat
- FOXP3 Variants Are Independently Associated With Transforming Growth Factor B1 Plasma Levels in Female Patients With Inflammatory Bowel DiseaseDokumen9 halamanFOXP3 Variants Are Independently Associated With Transforming Growth Factor B1 Plasma Levels in Female Patients With Inflammatory Bowel DiseasePaula KikuchiBelum ada peringkat
- Role of Imunity in Ap PDFDokumen9 halamanRole of Imunity in Ap PDFtanveer aliBelum ada peringkat
- Royal JellyDokumen10 halamanRoyal JellyninukBelum ada peringkat
- StatistikDokumen7 halamanStatistikAkhmad Ikhwan BaidlowiBelum ada peringkat
- Dexamethasone Inhibits IL-9 Production by Human T CellsDokumen9 halamanDexamethasone Inhibits IL-9 Production by Human T Cellsciara babyBelum ada peringkat
- Assessment of Splenic Function: ReviewDokumen9 halamanAssessment of Splenic Function: Reviewjosias_aragao3527Belum ada peringkat
- Proinflammatory Cytokines (IL-6, IL-8), Cytokine Inhibitors (IL-6sR, sTNFRII) and Anti-Inflammatory Cytokines (IL-10, IL-13) in The Pathogenesis of Sepsis in Newborns and InfantsDokumen6 halamanProinflammatory Cytokines (IL-6, IL-8), Cytokine Inhibitors (IL-6sR, sTNFRII) and Anti-Inflammatory Cytokines (IL-10, IL-13) in The Pathogenesis of Sepsis in Newborns and InfantsAstri Arri FebriantiBelum ada peringkat
- 1 SMDokumen4 halaman1 SMHanifa AdaniBelum ada peringkat
- tmp966C TMPDokumen10 halamantmp966C TMPFrontiersBelum ada peringkat
- Membranous Nephropathy: From Models To Man: Review SeriesDokumen8 halamanMembranous Nephropathy: From Models To Man: Review SeriesmmsBelum ada peringkat
- DoosDokumen10 halamanDoosmmsBelum ada peringkat
- Lupus Annals MI 2013Dokumen16 halamanLupus Annals MI 2013mmsBelum ada peringkat
- Renal Physiology For The Clinician Full Series CJASN PDFDokumen230 halamanRenal Physiology For The Clinician Full Series CJASN PDFmms100% (3)
- Molecular ImmunologyDokumen12 halamanMolecular ImmunologymmsBelum ada peringkat
- Cateteres Arteriaes, Central y Arteria PulmonarDokumen13 halamanCateteres Arteriaes, Central y Arteria PulmonarmmsBelum ada peringkat
- Goal-Directed Resuscitation For Patients With Early Septic ShockDokumen11 halamanGoal-Directed Resuscitation For Patients With Early Septic ShockmmsBelum ada peringkat
- Oncology S0360301616003242-MainDokumen20 halamanOncology S0360301616003242-Mainmms0% (1)
- 1 s2.0 S0026049515003650 MainDokumen12 halaman1 s2.0 S0026049515003650 MainmmsBelum ada peringkat
- Neuroscience LettersDokumen7 halamanNeuroscience LettersmmsBelum ada peringkat
- Intraabdominal InfectionsDokumen17 halamanIntraabdominal InfectionsmmsBelum ada peringkat
- 1 s2.0 S1878648013709881 MainDokumen1 halaman1 s2.0 S1878648013709881 MainmmsBelum ada peringkat
- Depresion en GeriatricosDokumen6 halamanDepresion en GeriatricosmmsBelum ada peringkat
- Hayek 2015Dokumen10 halamanHayek 2015mmsBelum ada peringkat
- CamScanner 10-29-2020 13.11.47Dokumen69 halamanCamScanner 10-29-2020 13.11.47Yuki ChanBelum ada peringkat
- Endocrine Organs 2004Dokumen323 halamanEndocrine Organs 2004Game TesterBelum ada peringkat
- Apex o GenesisDokumen54 halamanApex o GenesisbrahmannaBelum ada peringkat
- Anatomy AIIMSDokumen18 halamanAnatomy AIIMSvkBelum ada peringkat
- Yoga ExercisesDokumen10 halamanYoga ExercisesAna wulandariBelum ada peringkat
- Thyroid SwellingDokumen8 halamanThyroid SwellingDn Ezrinah Dn Esham50% (2)
- GastritisDokumen10 halamanGastritisMario100% (2)
- General Biology 1 NOTES PDFDokumen18 halamanGeneral Biology 1 NOTES PDFRose Mendizabal82% (17)
- Human Body EssayDokumen1 halamanHuman Body EssayAtlas CruxBelum ada peringkat
- A Study On Employee MotivationDokumen113 halamanA Study On Employee Motivationk eswari100% (2)
- Peptic Ulcers: DR Yotham Phiri Mmed (Surg)Dokumen46 halamanPeptic Ulcers: DR Yotham Phiri Mmed (Surg)Emmanuel MukukaBelum ada peringkat
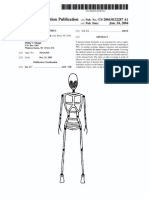
- Patente Endoesqueleto para Sex Doll SiliconaDokumen12 halamanPatente Endoesqueleto para Sex Doll SiliconaGabriel De Jesus100% (1)
- Final Exam Notes (Biology)Dokumen8 halamanFinal Exam Notes (Biology)410230675Belum ada peringkat
- Encoding: Cognitive Perspective: Fergus I. M. .CraikDokumen9 halamanEncoding: Cognitive Perspective: Fergus I. M. .Craikambar tri wahyuniBelum ada peringkat
- Care of Clients With Neurologic Deficit: By: Elmer G. Organia, RNDokumen233 halamanCare of Clients With Neurologic Deficit: By: Elmer G. Organia, RNTina TalmadgeBelum ada peringkat
- Aplastic Anemia Case Study FINALDokumen53 halamanAplastic Anemia Case Study FINALMatthew Rich Laguitao Cortes100% (6)
- Airway Management - IntubationDokumen31 halamanAirway Management - IntubationRajaBelum ada peringkat
- Shoku AnafilaktikDokumen16 halamanShoku AnafilaktikindeenikeBelum ada peringkat
- Isometric Power Revolution Part2Dokumen86 halamanIsometric Power Revolution Part2Smriti Ichopra100% (15)
- Complex Anorectal Disorders PDFDokumen817 halamanComplex Anorectal Disorders PDFGabriela Balan100% (2)
- 1.4 Selecton of Blood BagDokumen3 halaman1.4 Selecton of Blood BagBALAJIBelum ada peringkat
- Cancer Case StudyDokumen15 halamanCancer Case StudyRobin HaliliBelum ada peringkat
- ReceptoretDokumen37 halamanReceptoretBlodin ZylfiuBelum ada peringkat
- Toltec Body Healing EbookDokumen204 halamanToltec Body Healing EbookDanielle Grace100% (3)
- Anaphy Reviewer PrefinalsDokumen12 halamanAnaphy Reviewer PrefinalsNicole Faith L. NacarioBelum ada peringkat
- Atlas2Dokumen49 halamanAtlas2Andjela StojanovicBelum ada peringkat
- Laboratory Tests InterpretationDokumen12 halamanLaboratory Tests InterpretationKaloy Kamao100% (1)
- Cell Types SeDokumen16 halamanCell Types SeKendallBelum ada peringkat
- Uvmakalem PDFDokumen13 halamanUvmakalem PDFIsidoraVelimirovicBelum ada peringkat
- Group Case Study Obstetrics Group 5 From SMUDokumen84 halamanGroup Case Study Obstetrics Group 5 From SMUJosephine Mae TumanutBelum ada peringkat