Anda mungkin juga menyukai
- Evaluación de independencia funcional infantil (WEEFIMDokumen23 halamanEvaluación de independencia funcional infantil (WEEFIMMarisa De Luca100% (1)
- BursitisDokumen7 halamanBursitisgamers juarezBelum ada peringkat
- Antibióticos y bactericidas naturales para pecesDokumen13 halamanAntibióticos y bactericidas naturales para pecesJavier RodríguezBelum ada peringkat
- 0.7 Tejido CartilaginosoDokumen10 halaman0.7 Tejido CartilaginosoVale IsbarboBelum ada peringkat
- Metrado de Cantidades Iluminacion Exterior e InteriorDokumen6 halamanMetrado de Cantidades Iluminacion Exterior e InteriorOperaciones YJJBelum ada peringkat
- La apigenina, la kriptonita del cáncerDokumen3 halamanLa apigenina, la kriptonita del cáncertito cuadrosBelum ada peringkat
- 2608 2718 1 PBDokumen13 halaman2608 2718 1 PBChristiam Zamalloa Portocarrero100% (1)
- Cronograma de Talleres para El Fortalecimiento de La Vida UniversitariaDokumen5 halamanCronograma de Talleres para El Fortalecimiento de La Vida UniversitariaMiguel Alfonso VargasBelum ada peringkat
- Insuficiencia Renal Aguda y CronicaDokumen50 halamanInsuficiencia Renal Aguda y CronicaMiller ZeaBelum ada peringkat
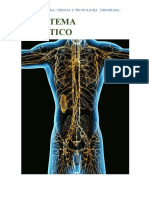
- Sistema linfático y sus funcionesDokumen7 halamanSistema linfático y sus funcionesdaniela mogollonBelum ada peringkat
- Atlas de Parasitología 2018Dokumen47 halamanAtlas de Parasitología 2018J 8aBelum ada peringkat
- Funciones del sistema respiratorioDokumen35 halamanFunciones del sistema respiratorioGabriel Zúñiga MartinezBelum ada peringkat
- Copia de Copia de Manual de PoesDokumen219 halamanCopia de Copia de Manual de PoesAdal Berrocales100% (1)
- 208 Evaluación de Tratamientos Químicos Más Fosfito de Calcio para El Control de Antracnosis (Ascochyta Pisi) en Cultivo de Arveja (Pisum Sativum L)Dokumen88 halaman208 Evaluación de Tratamientos Químicos Más Fosfito de Calcio para El Control de Antracnosis (Ascochyta Pisi) en Cultivo de Arveja (Pisum Sativum L)Wilmar Lizcano DuranBelum ada peringkat
- Tipos de metamorfosis insectos: hemimetabolismo vs holometabolismoDokumen1 halamanTipos de metamorfosis insectos: hemimetabolismo vs holometabolismorolinBelum ada peringkat
- BIpahuaaDokumen101 halamanBIpahuaaEdmundo68Belum ada peringkat
- Adaptacion CurricularDokumen4 halamanAdaptacion CurricularOmar Tallacagua LaimeBelum ada peringkat
- Via Piramidal ResumenDokumen9 halamanVia Piramidal ResumenJuan CastroBelum ada peringkat
- ArdiendoDokumen71 halamanArdiendo1999kauabx100% (1)
- Prueba Comprension Lectora 2 MedioDokumen5 halamanPrueba Comprension Lectora 2 MedioCar Los Daniel RamírezBelum ada peringkat
- Metaboliso Fungi2013avDokumen14 halamanMetaboliso Fungi2013avJuanBarbosaBelum ada peringkat
- FormulasDokumen7 halamanFormulasCami BattistelliBelum ada peringkat
- SenaDokumen19 halamanSenaKhiara DelgadoBelum ada peringkat
- Mapa Conceptual Enfoque AdministrativoDokumen1 halamanMapa Conceptual Enfoque AdministrativoEiner Zambrano100% (2)
- 135Dokumen143 halaman135rodorsBelum ada peringkat
- Glosario Biologia CelularDokumen34 halamanGlosario Biologia CelularGabriela DíazBelum ada peringkat
- Alimentos TransgenicosDokumen12 halamanAlimentos TransgenicosHernan MejiaBelum ada peringkat
- Diseña Tu Rutina de EntrenamientoDokumen15 halamanDiseña Tu Rutina de EntrenamientoHugo Aureliano Mora Arias100% (2)
- 669 Texto 669 1 10 20120719Dokumen15 halaman669 Texto 669 1 10 20120719ANGELO BENAVIDES GRANADOSBelum ada peringkat
- AnomalaseritrocitosDokumen5 halamanAnomalaseritrocitosYael Alejandro Garcia MarinBelum ada peringkat