Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Localization of MEG Human Brain Responses To Retinotopic Visual Stimuli With Contrasting Source Reconstruction ApproachesDokumen16 halamanLocalization of MEG Human Brain Responses To Retinotopic Visual Stimuli With Contrasting Source Reconstruction ApproachesnikadonBelum ada peringkat
- Individual Differences in Mental Imagery Tasks: A Study of Visual Thinkers and Verbal ThinkersDokumen8 halamanIndividual Differences in Mental Imagery Tasks: A Study of Visual Thinkers and Verbal ThinkersnikadonBelum ada peringkat
- Neuroimage: Justin Ales, Thom Carney, Stanley A. KleinDokumen9 halamanNeuroimage: Justin Ales, Thom Carney, Stanley A. KleinnikadonBelum ada peringkat
- fMRI Retinotopic Mapping-Step by StepDokumen19 halamanfMRI Retinotopic Mapping-Step by StepnikadonBelum ada peringkat
- Matrix Analysis, CAAM 335, Spring 2012: Steven J CoxDokumen98 halamanMatrix Analysis, CAAM 335, Spring 2012: Steven J CoxnikadonBelum ada peringkat
- Babel 2Dokumen217 halamanBabel 2nikadonBelum ada peringkat
- Concrete Centre SamplingDokumen1 halamanConcrete Centre SamplingEmir MuslićBelum ada peringkat
- Chapter 01Dokumen12 halamanChapter 01nikadonBelum ada peringkat
- Chapter 02Dokumen10 halamanChapter 02nikadonBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Assessment of The Use of A Dental Mouth MirrorDokumen7 halamanAssessment of The Use of A Dental Mouth MirrorIOSRjournalBelum ada peringkat
- AMCA Study Guide PhlebotomyDokumen34 halamanAMCA Study Guide Phlebotomyayoonia100% (1)
- Treat CMD Gnat NMD Chan 2003Dokumen32 halamanTreat CMD Gnat NMD Chan 2003Constantin IonutBelum ada peringkat
- Soap NotesDokumen4 halamanSoap NotesLatora Gardner Boswell100% (3)
- Behavior Neurobiology of Alcohol AddictionDokumen722 halamanBehavior Neurobiology of Alcohol AddictionDassaev Fritz100% (1)
- Cam4 6 1861Dokumen10 halamanCam4 6 1861Ćatke TkećaBelum ada peringkat
- No-Scar Transoral Thyroglossal Duct Cyst Excision in ChildrenDokumen21 halamanNo-Scar Transoral Thyroglossal Duct Cyst Excision in ChildrenMishel Rodriguez GuzmanBelum ada peringkat
- GMS 6871 Health Sciences Ethics Syllabus Summer 15Dokumen18 halamanGMS 6871 Health Sciences Ethics Syllabus Summer 15Ahmad AlSwaisBelum ada peringkat
- Green Power DistributionDokumen30 halamanGreen Power DistributionRazvan MarcuBelum ada peringkat
- Psych-K Practical ApplicationsDokumen10 halamanPsych-K Practical Applicationsdeluca4482% (17)
- Final Letter To NLM Re - IJOEH Nov 2017Dokumen35 halamanFinal Letter To NLM Re - IJOEH Nov 2017Celeste MonfortonBelum ada peringkat
- Guidance To Understanding Implications of IRMERDokumen52 halamanGuidance To Understanding Implications of IRMERrochimBelum ada peringkat
- Screening Questionnaire: Dha Eligibility LetterDokumen2 halamanScreening Questionnaire: Dha Eligibility LetterDr. Emad Elbadawy د عماد البدويBelum ada peringkat
- Speck - X-Ray Contrast Media - Overview, Use and Pharmaceutical AspectsDokumen134 halamanSpeck - X-Ray Contrast Media - Overview, Use and Pharmaceutical AspectsSimona Mariana DutuBelum ada peringkat
- Textbook of Forensic Medicine & Toxicology (VV Pillay)Dokumen513 halamanTextbook of Forensic Medicine & Toxicology (VV Pillay)s.misbaunnisa26100% (3)
- NCP For Ineffective Airway Clearance.Dokumen2 halamanNCP For Ineffective Airway Clearance.Vanessa Joy Contreras100% (1)
- 080 - Hospital Information Document For New StaffDokumen2 halaman080 - Hospital Information Document For New StaffleicesterbugBelum ada peringkat
- Asperger Syndrome in ChildrenDokumen8 halamanAsperger Syndrome in Childrenmaria_kazaBelum ada peringkat
- Goljan Errata SheetDokumen11 halamanGoljan Errata SheetVishala MishraBelum ada peringkat
- Non-Suicidal Self-Injury (NSSI) and Suicide: Criteria DifferentiationDokumen7 halamanNon-Suicidal Self-Injury (NSSI) and Suicide: Criteria DifferentiationTututWidyaNurAnggrainiBelum ada peringkat
- Paroxysmal Nocturnal Hemoglobinuria: Section 13Dokumen2 halamanParoxysmal Nocturnal Hemoglobinuria: Section 13MANGBelum ada peringkat
- LMC ICD-10 PowerPointDokumen77 halamanLMC ICD-10 PowerPointNicholas HenryBelum ada peringkat
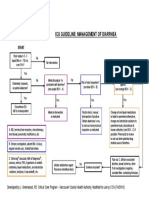
- Icu Guideline: Management of Diarrhea: StartDokumen1 halamanIcu Guideline: Management of Diarrhea: StartGracia VionaBelum ada peringkat
- Anaphy (Lab) ReviewerDokumen3 halamanAnaphy (Lab) ReviewerAgatha Cristie AndradaBelum ada peringkat
- Capstone OutlineDokumen3 halamanCapstone Outlineapi-395468231Belum ada peringkat
- Twin Block Appliance ThesisDokumen6 halamanTwin Block Appliance Thesisjessicahillnewyork100% (2)
- Report CapsicumDokumen105 halamanReport CapsicumRina WijayantiBelum ada peringkat
- 2.scope of Diagnostic RadiologyDokumen28 halaman2.scope of Diagnostic RadiologyphilipBelum ada peringkat
- Forensic Pharmacy & Forensic Pharmacist: Presented By: Ibrahim Shafqat Uzair Abid Fahad Wadood Shoiab ShakeelDokumen16 halamanForensic Pharmacy & Forensic Pharmacist: Presented By: Ibrahim Shafqat Uzair Abid Fahad Wadood Shoiab ShakeelJaved IqbalBelum ada peringkat
- Vaginal Discharge in DogsDokumen5 halamanVaginal Discharge in DogsDarmawan Prastyo100% (1)