Anda mungkin juga menyukai
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Three Kingdoms RevisitedDokumen152 halamanThree Kingdoms RevisitedBobbyGunarsoBelum ada peringkat
- À Å° Ó Ðåä - Sonnet of FountainDokumen5 halamanÀ Å° Ó Ðåä - Sonnet of FountainBobbyGunarsoBelum ada peringkat
- I Believe in Springtime - Rutter - EnglishDokumen3 halamanI Believe in Springtime - Rutter - EnglishBobbyGunarsoBelum ada peringkat
- Cardiac Pacemakers and Defibrillators 2nd Ed - C. Love (Landes, 2006) WW PDFDokumen183 halamanCardiac Pacemakers and Defibrillators 2nd Ed - C. Love (Landes, 2006) WW PDFJozsa Attila100% (1)
- Caring For Injured Reptiles2004Dokumen40 halamanCaring For Injured Reptiles2004SujayJainBelum ada peringkat
- Burt and Eklund's Dentistry, Dental Practice, and The Community, 7th EditionDokumen330 halamanBurt and Eklund's Dentistry, Dental Practice, and The Community, 7th EditionPhuong Thao75% (4)
- Metrics and Evaluation: Metric Sample Calculation Advantages DisadvantagesDokumen10 halamanMetrics and Evaluation: Metric Sample Calculation Advantages DisadvantagesBobbyGunarsoBelum ada peringkat
- Grade 8 Health Summative Test Alcohol and CigaretteDokumen3 halamanGrade 8 Health Summative Test Alcohol and CigaretteAlbert Ian Casuga100% (1)
- NCP OsteosarcomaDokumen6 halamanNCP OsteosarcomaNiksBelum ada peringkat
- PNLE Reviewer With Rationale 1Dokumen12 halamanPNLE Reviewer With Rationale 1Zyla Malinao100% (1)
- NCP For PTSDDokumen3 halamanNCP For PTSDjhaze09100% (4)
- Kirim Ukai HipertensiDokumen18 halamanKirim Ukai HipertensiBobbyGunarsoBelum ada peringkat
- Domiciliary Midwifery RDokumen16 halamanDomiciliary Midwifery Rswillymadhu83% (6)
- Persuasive Speech Outline - Marijauna FinalDokumen4 halamanPersuasive Speech Outline - Marijauna Finalapi-34805003388% (17)
- Wanda Wajang Purwa R Soetrisno 2 27Dokumen13 halamanWanda Wajang Purwa R Soetrisno 2 27BobbyGunarsoBelum ada peringkat
- 9781603841948Dokumen1 halaman9781603841948BobbyGunarsoBelum ada peringkat
- Kesehatan Dan Keadilan Sosial inDokumen20 halamanKesehatan Dan Keadilan Sosial inBobbyGunarsoBelum ada peringkat
- Sebuah Patung Singa Dari Candi Prambanan Diapit Dengan Dua Kapataru Dan KinaraDokumen28 halamanSebuah Patung Singa Dari Candi Prambanan Diapit Dengan Dua Kapataru Dan KinaraBobbyGunarsoBelum ada peringkat
- Barata Yudha Sunardi DMDokumen72 halamanBarata Yudha Sunardi DMBobbyGunarsoBelum ada peringkat
- Modul Matrikulasi FTSF - Bu BetiDokumen146 halamanModul Matrikulasi FTSF - Bu BetiBobbyGunarsoBelum ada peringkat
- Development of Cell-Cycle Inhibitors For Cancer Therapy: M.A. Dickson and G.K. SchwartzDokumen8 halamanDevelopment of Cell-Cycle Inhibitors For Cancer Therapy: M.A. Dickson and G.K. SchwartzBobbyGunarsoBelum ada peringkat
- Modul UKAI 2015 FarmakokimiaDokumen41 halamanModul UKAI 2015 FarmakokimiaBobbyGunarsoBelum ada peringkat
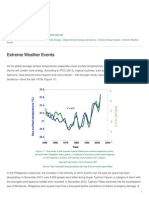
- Healing Earth - Extreme Weather Events - 2015-04-21Dokumen2 halamanHealing Earth - Extreme Weather Events - 2015-04-21BobbyGunarsoBelum ada peringkat
- Indonesia Raya For Symphony OrchestraDokumen13 halamanIndonesia Raya For Symphony OrchestraBobbyGunarsoBelum ada peringkat
- Indonesia Raya For PianoDokumen5 halamanIndonesia Raya For PianoBobbyGunarso50% (2)
- Water Density: Published On Healing EarthDokumen2 halamanWater Density: Published On Healing EarthBobbyGunarsoBelum ada peringkat
- Pasteur Eng MiddleDokumen4 halamanPasteur Eng MiddleRoykoBelum ada peringkat
- Inferior Vena Cava and Its TributariesDokumen12 halamanInferior Vena Cava and Its TributariesRahaf MajadlyBelum ada peringkat
- Chapter 3 Animal Tissues SolutionDokumen6 halamanChapter 3 Animal Tissues SolutionLucky ErrojuBelum ada peringkat
- DR Antonious CV N & AEDokumen27 halamanDR Antonious CV N & AEdoctorantoniBelum ada peringkat
- (Dental Discussion 1) - Hesy ReDokumen32 halaman(Dental Discussion 1) - Hesy ReethanBelum ada peringkat
- Chorti DictionaryDokumen285 halamanChorti DictionaryLuis Bedoya100% (1)
- Europe Heatwave: Why Are Temperatures On The Continent Soaring?Dokumen3 halamanEurope Heatwave: Why Are Temperatures On The Continent Soaring?Alec LoaiBelum ada peringkat
- Subacute and Chronic MeningitisDokumen8 halamanSubacute and Chronic MeningitisLucky PuspitasariBelum ada peringkat
- Magnesium Sulphate InjDokumen2 halamanMagnesium Sulphate InjmahgadBelum ada peringkat
- GI Bleeding in Patients Receiving Antiplatelets and Anticoagulant TherapyDokumen11 halamanGI Bleeding in Patients Receiving Antiplatelets and Anticoagulant TherapyTony LeeBelum ada peringkat
- Aloe VeraDokumen8 halamanAloe VeraOmar M SalahBelum ada peringkat
- Oxi Ferm Pluri-Test 78620Dokumen6 halamanOxi Ferm Pluri-Test 78620Ronny Miranda MaciasBelum ada peringkat
- Bio TherapyDokumen18 halamanBio TherapyChino RamirezBelum ada peringkat
- Application Form: Tata Aia Life Insurance Company LimitedDokumen6 halamanApplication Form: Tata Aia Life Insurance Company LimitedpritamBelum ada peringkat
- Allergic RhinitisDokumen8 halamanAllergic RhinitisJuanBelum ada peringkat
- Jewish Genetic DiseasesDokumen2 halamanJewish Genetic DiseasesReformJudaismBelum ada peringkat
- 2.1 (I) Know The Properties of Gas Exchange Surfaces in Living Organisms 2Dokumen16 halaman2.1 (I) Know The Properties of Gas Exchange Surfaces in Living Organisms 2YEUNG WAN CHUEN ANGELIQUEBelum ada peringkat
- DarwinDokumen118 halamanDarwinmrthumb100% (2)
- Changes in Diet and Lifestyle and Long-Term Weight Gain in Women and MenDokumen13 halamanChanges in Diet and Lifestyle and Long-Term Weight Gain in Women and MenemanvitoriaBelum ada peringkat
- لقطة شاشة 2023-02-02 في 11.30.55 ص PDFDokumen50 halamanلقطة شاشة 2023-02-02 في 11.30.55 ص PDFFaisal AlhiallBelum ada peringkat
- Colon Cancer Preventable, Detectable With ScreeningDokumen1 halamanColon Cancer Preventable, Detectable With ScreeningAnonymous s40aYyBelum ada peringkat