Anda mungkin juga menyukai
- Micosis Superficial Dra Eva GaragorriDokumen165 halamanMicosis Superficial Dra Eva GaragorriLuis Yehsuah Moran Amaya100% (1)
- Histologia CompletoDokumen91 halamanHistologia CompletoLuis Yehsuah Moran AmayaBelum ada peringkat
- Histologia CompletoDokumen91 halamanHistologia CompletoLuis Yehsuah Moran AmayaBelum ada peringkat
- TROFOBLASTODokumen16 halamanTROFOBLASTOArmando Chris Q67% (3)
- Parcial Endocrino4Dokumen18 halamanParcial Endocrino4Bianca CerecedesBelum ada peringkat
- Desarrollo embrionario de la 3ra a la 8va semanaDokumen7 halamanDesarrollo embrionario de la 3ra a la 8va semanaᴛᴡᴏ5Belum ada peringkat
- El Adn Como Base de La Vida - Semana 7 de Biologia 2doDokumen19 halamanEl Adn Como Base de La Vida - Semana 7 de Biologia 2doAbigailtulcanBelum ada peringkat
- Morfología Del Eritron Grupo #4Dokumen37 halamanMorfología Del Eritron Grupo #4Michelle Mejia FamiliaBelum ada peringkat
- Neoplasias Parte 1 Conceptos Generales y NomenclaturaDokumen14 halamanNeoplasias Parte 1 Conceptos Generales y NomenclaturaMARCOABADBelum ada peringkat
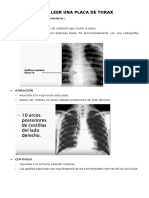
- Como Leer Una Placa de Torax y Estructuras Basicas AnatomicasDokumen23 halamanComo Leer Una Placa de Torax y Estructuras Basicas AnatomicasLuis Yehsuah Moran Amaya100% (1)
- DR Veller E. HipertensivaDokumen7 halamanDR Veller E. HipertensivaMiguel Badel100% (2)
- Banco de Preguntas II UnidadDokumen9 halamanBanco de Preguntas II UnidadOrlando Jesús Oruna DelgadoBelum ada peringkat
- EJERCICIO CALIFICADO Sesion 2 CENTENO PARARIDokumen5 halamanEJERCICIO CALIFICADO Sesion 2 CENTENO PARARITony Nouh100% (1)
- Vascularización e Inervación Del DorsoDokumen2 halamanVascularización e Inervación Del DorsoHector Javier Perez Martinez100% (2)
- Embriologia de Las ExtremidadesDokumen53 halamanEmbriologia de Las ExtremidadesAbel Vergara100% (1)
- Desarrollo de Las ExtremidadesDokumen35 halamanDesarrollo de Las ExtremidadesJorge Luis Del Aguila Hernandez50% (2)
- REGULACIÓN MOLECULAR DEL DESARROLLO CARDIACODokumen3 halamanREGULACIÓN MOLECULAR DEL DESARROLLO CARDIACOCarolina AquinoBelum ada peringkat
- Derivados de La Capa Germinativa EctodérmicaDokumen10 halamanDerivados de La Capa Germinativa EctodérmicaCamilo LunaBelum ada peringkat
- Guía de Práctica 12 2023Dokumen13 halamanGuía de Práctica 12 2023JAROLL JHANPOLL CUZCO MENDOZABelum ada peringkat
- Embriologia ALAN Esqueleto ApendicularDokumen14 halamanEmbriologia ALAN Esqueleto ApendicularAlan Ch Flores100% (3)
- Observación celularDokumen10 halamanObservación celularjuanBelum ada peringkat
- Guia Práctica 06Dokumen13 halamanGuia Práctica 06ZIUGASBelum ada peringkat
- Músculos EmbriologíaDokumen14 halamanMúsculos EmbriologíaAlicia Sigcho0% (1)
- MIELOMENIGOCELEDokumen16 halamanMIELOMENIGOCELEANTONELLA ABIGAIL GONZALEZ NARVAEZBelum ada peringkat
- Practica Semana 05 Reticulo EndoplasmaticoDokumen9 halamanPractica Semana 05 Reticulo EndoplasmaticoJeni Campos RamosBelum ada peringkat
- Mesos, Celoma y sus anomalíasDokumen5 halamanMesos, Celoma y sus anomalíasAdolfo Antonio Ríos AlcortaBelum ada peringkat
- Guía Pract 4ta SesiónDokumen8 halamanGuía Pract 4ta SesiónFatima Palacios GeronimoBelum ada peringkat
- Fisiopatologia Sindrome de LeighDokumen8 halamanFisiopatologia Sindrome de Leighgrupo trabajo inmuno 2017Belum ada peringkat
- Aponeurosis de La Region Antero Lateral Del ToraxDokumen3 halamanAponeurosis de La Region Antero Lateral Del ToraxJorge Cornelio Mainato Shagñay0% (1)
- MeiosisDokumen13 halamanMeiosisJAROLL JHANPOLL CUZCO MENDOZA100% (1)
- Circulación fetal y neonatalDokumen3 halamanCirculación fetal y neonatalAlfonso Castro100% (3)
- Embriología 6Dokumen25 halamanEmbriología 6Nicole DíazBelum ada peringkat
- Informe #7Dokumen3 halamanInforme #7Gabriel FreireBelum ada peringkat
- 12 Estimación de Costos y Administracion de PyDokumen35 halaman12 Estimación de Costos y Administracion de PyYOMIDES LEONCIO LOLOY CAPABelum ada peringkat
- Práctica #14: Michael Kevin Mejia TarazonaDokumen20 halamanPráctica #14: Michael Kevin Mejia TarazonaZIUGASBelum ada peringkat
- Fisiopatologia Del Sistema RespiratorioDokumen39 halamanFisiopatologia Del Sistema RespiratorioMayra Milagros Ruiz SanchezBelum ada peringkat
- Regulación Molecular Del Desarrollo CardíacoDokumen4 halamanRegulación Molecular Del Desarrollo CardíacoDar Muñoz0% (1)
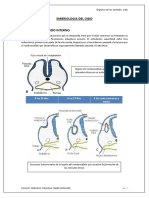
- Embriologia Del OidoDokumen21 halamanEmbriologia Del OidoNadiaMossanoBelum ada peringkat
- SomitasDokumen4 halamanSomitasyacson cano valenciaBelum ada peringkat
- Paralisis en La HemicaraDokumen20 halamanParalisis en La HemicaraAlejandro Zuñiga100% (1)
- Tema 2 BioenergeticaDokumen17 halamanTema 2 BioenergeticaCristhian Macalupú Arismendiz100% (1)
- Embriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMDokumen83 halamanEmbriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMCarlos GonzalesBelum ada peringkat
- Cavidades CorporalesDokumen4 halamanCavidades CorporalesLana KluyBelum ada peringkat
- GinecologiaDokumen62 halamanGinecologiaLuis Miguel AjilaBelum ada peringkat
- Anemias HarrisonDokumen9 halamanAnemias HarrisonAndrea GallegoBelum ada peringkat
- Cuestionario Embriologia IIBDokumen64 halamanCuestionario Embriologia IIBJim SchoBelum ada peringkat
- Estructura y función del sacro y cóccixDokumen13 halamanEstructura y función del sacro y cóccixmaria fernandaBelum ada peringkat
- Ejercicios Capitulo 7Dokumen5 halamanEjercicios Capitulo 7julian0% (1)
- Cuál Es La Importancia Del Núcleo CelularDokumen2 halamanCuál Es La Importancia Del Núcleo CelularManuela Arcila Santa MariaBelum ada peringkat
- Práctica 4 - Tejido Epitelial y MuscularDokumen12 halamanPráctica 4 - Tejido Epitelial y MuscularYESEIDY DAHIANA ZUTA SANTILL�NBelum ada peringkat
- EmbriologíaDokumen34 halamanEmbriologíaJosehanny NGBelum ada peringkat
- Anatomía - Práctica Sesión 07Dokumen4 halamanAnatomía - Práctica Sesión 07Kenyu Esquivel MendozaBelum ada peringkat
- SomitasDokumen4 halamanSomitasMayra AlejandraBelum ada peringkat
- Vías de Transducción Intracelular de SeñalesDokumen8 halamanVías de Transducción Intracelular de SeñalesCristina PazmiñoBelum ada peringkat
- s11 Lectura Laboratorio 10Dokumen22 halamans11 Lectura Laboratorio 10Oriana MassielBelum ada peringkat
- Precursores Del Cáncer Gástrico: Factores de Riesgo, Condiciones y Lesiones Premalignas de La Mucosa GástricaDokumen6 halamanPrecursores Del Cáncer Gástrico: Factores de Riesgo, Condiciones y Lesiones Premalignas de La Mucosa GástricaRIsas TravesurasBelum ada peringkat
- Tuberculosis causas síntomasDokumen2 halamanTuberculosis causas síntomasFotocopias LumaBelum ada peringkat
- Documento Sin TítuloDokumen10 halamanDocumento Sin TítuloHania Clavijo100% (1)
- Resumen - TraqueaDokumen4 halamanResumen - TraqueaLucas pperesBelum ada peringkat
- Guía Pract 8va SesiónDokumen4 halamanGuía Pract 8va SesiónEvelyn LMBelum ada peringkat
- Anomalias Del Conducto VitelinoDokumen23 halamanAnomalias Del Conducto VitelinoKaren Campos ManjaresBelum ada peringkat
- Banco Preguntas PlacentaDokumen1 halamanBanco Preguntas PlacentaEstefanía RiveraBelum ada peringkat
- Estructura y relaciones anatómicas del intestino delgadoDokumen3 halamanEstructura y relaciones anatómicas del intestino delgadoJuan Perez67% (3)
- Region CarotideaDokumen9 halamanRegion CarotideaDiego Vazquez ChaireBelum ada peringkat
- Situación - Problematica DOCENTEDokumen2 halamanSituación - Problematica DOCENTENayara MirellyBelum ada peringkat
- Problemas de BioelectricidadDokumen1 halamanProblemas de BioelectricidadOmar Benjamin FDBelum ada peringkat
- Diversidad Étnica y Lingüística en El PerúDokumen2 halamanDiversidad Étnica y Lingüística en El Perúmalmavalero0% (1)
- Cuestionario Biol Del Desar 2020 JunioDokumen10 halamanCuestionario Biol Del Desar 2020 JunioMvz Franklin MamaniBelum ada peringkat
- Neuropatias 2019.Ppt - DegrabadaDokumen108 halamanNeuropatias 2019.Ppt - DegrabadaLuis Yehsuah Moran AmayaBelum ada peringkat
- Genes Del DesarrolloDokumen97 halamanGenes Del DesarrolloLuis Yehsuah Moran AmayaBelum ada peringkat
- Psicological Studies and RecommendationsDokumen3 halamanPsicological Studies and RecommendationsLuis Yehsuah Moran AmayaBelum ada peringkat
- 7Dokumen48 halaman7Luis Yehsuah Moran AmayaBelum ada peringkat
- Psicological Studies and RecommendationsDokumen3 halamanPsicological Studies and RecommendationsLuis Yehsuah Moran AmayaBelum ada peringkat
- EMBRIOLOGIADokumen21 halamanEMBRIOLOGIALuis Yehsuah Moran AmayaBelum ada peringkat
- Histología de la PielDokumen20 halamanHistología de la PielLuis Yehsuah Moran AmayaBelum ada peringkat
- SistemaDokumen79 halamanSistemaLuis Yehsuah Moran AmayaBelum ada peringkat
- Presión Venosa Yugular y Trazo YugularDokumen3 halamanPresión Venosa Yugular y Trazo YugularLuis Yehsuah Moran AmayaBelum ada peringkat
- Andrea Tooley Study TechniquesDokumen5 halamanAndrea Tooley Study TechniquesLuis Yehsuah Moran AmayaBelum ada peringkat
- Practica Final 2017Dokumen1 halamanPractica Final 2017Luis Yehsuah Moran AmayaBelum ada peringkat
- Seminario farmacología principales fármacos revisarDokumen3 halamanSeminario farmacología principales fármacos revisarcrt20003Belum ada peringkat
- Digestion de CarbohidratosDokumen12 halamanDigestion de CarbohidratosLuis Yehsuah Moran AmayaBelum ada peringkat
- Funciones Y Clasificación de Las Células Dendríticas: F. R - P, P.J. S C, M.A. R, M. P, V. M, E. R - V, J.C. G - VDokumen25 halamanFunciones Y Clasificación de Las Células Dendríticas: F. R - P, P.J. S C, M.A. R, M. P, V. M, E. R - V, J.C. G - Vandrey1232Belum ada peringkat
- MAPADokumen1 halamanMAPALuis Yehsuah Moran AmayaBelum ada peringkat
- Caso Clinico Fisiopato Acv FinalDokumen15 halamanCaso Clinico Fisiopato Acv FinalLuis Yehsuah Moran AmayaBelum ada peringkat
- Funciones Y Clasificación de Las Células Dendríticas: F. R - P, P.J. S C, M.A. R, M. P, V. M, E. R - V, J.C. G - VDokumen25 halamanFunciones Y Clasificación de Las Células Dendríticas: F. R - P, P.J. S C, M.A. R, M. P, V. M, E. R - V, J.C. G - Vandrey1232Belum ada peringkat
- SPSS BDDokumen21 halamanSPSS BDLuis Yehsuah Moran AmayaBelum ada peringkat
- Repaso HistologíaDokumen53 halamanRepaso HistologíaLuis Yehsuah Moran AmayaBelum ada peringkat
- Articulo Interesante Checar para Expo PDFDokumen16 halamanArticulo Interesante Checar para Expo PDFLuis Yehsuah Moran AmayaBelum ada peringkat
- Consentimiento InformadoDokumen1 halamanConsentimiento InformadoLuis Yehsuah Moran AmayaBelum ada peringkat
- Gestion de Riesgos, ManualDokumen43 halamanGestion de Riesgos, ManualFidel Montes GodoyBelum ada peringkat
- Guillen Barre Zika PDFDokumen8 halamanGuillen Barre Zika PDFLuis Yehsuah Moran AmayaBelum ada peringkat
- Anomalías CromosómicasDokumen12 halamanAnomalías CromosómicasMaríaCristinaVásquezBelum ada peringkat
- Corpúsculo de BarrDokumen3 halamanCorpúsculo de BarrMaria Teresa Rosal GarciaBelum ada peringkat
- Tejido EpiterialDokumen11 halamanTejido EpiterialNene Zavala PalmaBelum ada peringkat
- Componentes Sanguíneos y CoagulaciónDokumen3 halamanComponentes Sanguíneos y CoagulaciónCitlali Sarai Soto ReyesBelum ada peringkat
- Estructura bacteriana taller 2 Dr. ArochaDokumen92 halamanEstructura bacteriana taller 2 Dr. ArochaJohnny GarrilloBelum ada peringkat
- Multiple Choice Biologia CBCDokumen9 halamanMultiple Choice Biologia CBCjochuchacon10Belum ada peringkat
- UNIDAD III - La Celula Como La Unidad de Los Seres VivosDokumen60 halamanUNIDAD III - La Celula Como La Unidad de Los Seres VivosEmmanuel PaulinoBelum ada peringkat
- Virus Características GeneralesDokumen4 halamanVirus Características GeneralesSamuel MartinezBelum ada peringkat
- Sintesis de Acidos GrasosDokumen12 halamanSintesis de Acidos GrasosKARLA VERONICA DIAZ AMAYABelum ada peringkat
- Introduccion - EpitelialDokumen5 halamanIntroduccion - EpitelialNicolas GomezBelum ada peringkat
- Desarrollo VellosidadesDokumen9 halamanDesarrollo VellosidadesMagic_OverBelum ada peringkat
- Cultivos celulares: historia, tipos y líneas celularesDokumen23 halamanCultivos celulares: historia, tipos y líneas celularesYuli Miers0% (1)
- Sarcodinos ComensalesDokumen26 halamanSarcodinos ComensalesMiguel RivadeneiraBelum ada peringkat
- HISTOCOMPATIBILIDADDokumen24 halamanHISTOCOMPATIBILIDADRaquel Lopez AndresBelum ada peringkat
- Cuadro Resumen Código GenéticoDokumen1 halamanCuadro Resumen Código GenéticojuliaddazaBelum ada peringkat
- Análisis de laboratorio con resultados de química sanguínea, hematología y coagulaciónDokumen2 halamanAnálisis de laboratorio con resultados de química sanguínea, hematología y coagulaciónJulián Andrés PosadaBelum ada peringkat
- Esquema Sobre Las Causas Del FLAPNSDokumen1 halamanEsquema Sobre Las Causas Del FLAPNSDomenica ErazoBelum ada peringkat
- Funciones proteínicasDokumen29 halamanFunciones proteínicasMildred Andrea Olguin GonzalezBelum ada peringkat
- Actividad 1 Laboratorio Virtual 3roDokumen2 halamanActividad 1 Laboratorio Virtual 3roJuan ParraBelum ada peringkat
- MESOTERAPIA Dr. Villalba VzlaDokumen67 halamanMESOTERAPIA Dr. Villalba VzlaCarlos CarrenoBelum ada peringkat
- Fisiología: Sesión #5: Sinapsis: Vereau Gutierrez, Miguel Angel FroilanDokumen35 halamanFisiología: Sesión #5: Sinapsis: Vereau Gutierrez, Miguel Angel FroilanSebastian Miguel Ramos ToledoBelum ada peringkat
- Nutricion HeterotofaDokumen2 halamanNutricion HeterotofaMarcela RuizBelum ada peringkat
- Mapa Mental de La Ce 155135 Downloadable 4247210Dokumen2 halamanMapa Mental de La Ce 155135 Downloadable 4247210marcoscastellanosvalverde2004Belum ada peringkat
- 01 GAMETOGÉNESIS IIIIb OOGÉNESISDokumen30 halaman01 GAMETOGÉNESIS IIIIb OOGÉNESISAdolfo Antonio Ríos AlcortaBelum ada peringkat
- Mm. Cuestionario Bacteriología Básica IDokumen5 halamanMm. Cuestionario Bacteriología Básica IDaniela Guerrero.Belum ada peringkat
- REACCIONES ENZIMÁTICAS DE OXIDO-REDUCCIÓNDokumen1 halamanREACCIONES ENZIMÁTICAS DE OXIDO-REDUCCIÓNMoises Hernandes100% (1)
- AcetilcolinaDokumen2 halamanAcetilcolinaPRISCILA YAMILET JIMENEZ MANZOBelum ada peringkat
- 4-Citometria de FlujoDokumen41 halaman4-Citometria de FlujoTatiana MendozaBelum ada peringkat
- Cuestionario de Tejido ConectivoDokumen4 halamanCuestionario de Tejido ConectivoDiana AgipBelum ada peringkat