Anda mungkin juga menyukai
- Distribucion de Una Planta de ProduccionDokumen2 halamanDistribucion de Una Planta de ProduccionNelly Johanna Soto CastañedaBelum ada peringkat
- Ensayo Importancia AICDokumen1 halamanEnsayo Importancia AICNelly Johanna Soto CastañedaBelum ada peringkat
- Estudio de Caso - Semana 4Dokumen2 halamanEstudio de Caso - Semana 4Nelly Johanna Soto Castañeda83% (30)
- Nelly Johanna Soto CastañedaDokumen10 halamanNelly Johanna Soto CastañedaNelly Johanna Soto CastañedaBelum ada peringkat
- Informe EjecutivoDokumen3 halamanInforme EjecutivoNelly Johanna Soto CastañedaBelum ada peringkat
- Nosotras Morimos para Que Ustedes Puedan Tener Sus SmartphonesDokumen3 halamanNosotras Morimos para Que Ustedes Puedan Tener Sus SmartphonesNelly Johanna Soto CastañedaBelum ada peringkat
- Oración de RecomendaciónDokumen6 halamanOración de RecomendaciónNelly Johanna Soto CastañedaBelum ada peringkat
- Biodiesel de Producción Por Un Proceso Continuo Utilizando Un Catalizador HeterogeneoDokumen6 halamanBiodiesel de Producción Por Un Proceso Continuo Utilizando Un Catalizador HeterogeneoNelly Johanna Soto Castañeda100% (1)
- Planta de Produccion de MCVDokumen12 halamanPlanta de Produccion de MCVNelly Johanna Soto CastañedaBelum ada peringkat
- Escalado de Reactores QuimicosDokumen18 halamanEscalado de Reactores QuimicosNelly Johanna Soto Castañeda100% (1)
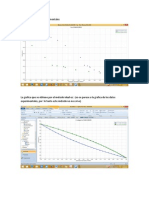
- Grafica de Los Datos ExperimentalesDokumen4 halamanGrafica de Los Datos ExperimentalesNelly Johanna Soto CastañedaBelum ada peringkat
- Prospecto Urzac I0058-4Dokumen1 halamanProspecto Urzac I0058-4Harold83Belum ada peringkat
- TorchDokumen40 halamanTorchTony AquinoBelum ada peringkat
- Sistema EndocrinoDokumen2 halamanSistema Endocrinoluis peraltaBelum ada peringkat
- Tratamiento de La Ansiedad GeneralizadaDokumen16 halamanTratamiento de La Ansiedad GeneralizadaAndy Soles VillanuevaBelum ada peringkat
- VACUNACIÓN EN EL RECIÉN NACIDO PREMATURO y EmbarazoDokumen5 halamanVACUNACIÓN EN EL RECIÉN NACIDO PREMATURO y EmbarazoCesar PinachoBelum ada peringkat
- Calzado PlantillasDokumen30 halamanCalzado PlantillasAlejandro Folch100% (5)
- Trichosporon BeigeliiDokumen11 halamanTrichosporon BeigeliiADRIAN0% (1)
- Los Lisosomas 3Dokumen16 halamanLos Lisosomas 3wilber rojasBelum ada peringkat
- Los Anticuerpos Anti EritrociticosDokumen15 halamanLos Anticuerpos Anti Eritrociticosfaster69Belum ada peringkat
- BRIXOMETRODokumen37 halamanBRIXOMETROJorge Chavez QuispeBelum ada peringkat
- Las Fantasías y El Acompañamiento TerapéuticoDokumen6 halamanLas Fantasías y El Acompañamiento Terapéuticosalome0% (1)
- Plan de Trabajo Estrategia TuberculosisDokumen5 halamanPlan de Trabajo Estrategia Tuberculosiscrlos medBelum ada peringkat
- WWe 4 VN JWB DDokumen166 halamanWWe 4 VN JWB DLorena Guadalupe Terreros BucioBelum ada peringkat
- Sindrome Antifosfolipidico y EmbarazoDokumen43 halamanSindrome Antifosfolipidico y EmbarazoGladyola Pulgarito0% (1)
- Sesión 12Dokumen8 halamanSesión 12Silvia Monica VillanuevaBelum ada peringkat
- Las EmosionesDokumen4 halamanLas EmosionesElizabeth ES100% (1)
- Ensayo Final Introducción Al Análisis InstrumentalDokumen6 halamanEnsayo Final Introducción Al Análisis InstrumentalJuan Garzón LamarqueBelum ada peringkat
- Ctom Evaluacion PsicologicaDokumen7 halamanCtom Evaluacion PsicologicaveronicafinoliBelum ada peringkat
- Kineflaxia para FisioterapeDokumen2 halamanKineflaxia para FisioterapeMariia Jose BurgosBelum ada peringkat
- Clasificación de La AscitisDokumen8 halamanClasificación de La AscitisAnnelli MichelleBelum ada peringkat
- A B C TraumaDokumen38 halamanA B C Traumadaniel camuBelum ada peringkat
- Epilepsia Completo PDFDokumen16 halamanEpilepsia Completo PDFBiol. Miguel Angel Gutiérrez Domínguez100% (1)
- La Célula Como Unidad de Salud y EnfermedadDokumen55 halamanLa Célula Como Unidad de Salud y EnfermedadJoselline Alva Encinas100% (3)
- MonografiaDokumen20 halamanMonografiaDavid Piscoya SeguraBelum ada peringkat
- S 8 SaponinasDokumen16 halamanS 8 SaponinasRocio Reyes Cueva100% (2)
- Medicina AyurvedicaDokumen5 halamanMedicina AyurvedicajorgebreaBelum ada peringkat
- Aprendizaje Basado en ProblemasDokumen7 halamanAprendizaje Basado en ProblemasPedro LorancaBelum ada peringkat
- Trastornos TraumatologicosDokumen21 halamanTrastornos TraumatologicosHeidy Marisa Morales CarttyBelum ada peringkat
- Apuntes ServicioCuidadosCrÃ-ticosNeonato Arial 8Â Ed - 2 Abril-2015 OKDokumen300 halamanApuntes ServicioCuidadosCrÃ-ticosNeonato Arial 8Â Ed - 2 Abril-2015 OKJuan Lujan Salas100% (1)
- Libro de Anatomia Patologica M5Dokumen0 halamanLibro de Anatomia Patologica M5Zoila Gutierrez AyalaBelum ada peringkat