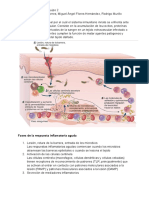
Anda mungkin juga menyukai
- La Preocupación Inútil - Laura Vera PatierDokumen142 halamanLa Preocupación Inútil - Laura Vera PatierAna Barreiro88% (8)
- Semiología de Las Enfermedades CardiovascularesDokumen384 halamanSemiología de Las Enfermedades Cardiovascularesdcicare100% (3)
- Glándulas SuprarrenalesDokumen33 halamanGlándulas SuprarrenalesAngie SaileBelum ada peringkat
- Atlas-De-Bolsillo-De-Fisiologia-Humana. Silbernagl. Despopoulos PDFDokumen227 halamanAtlas-De-Bolsillo-De-Fisiologia-Humana. Silbernagl. Despopoulos PDFMauricio Zúñiga100% (1)
- Compuestos NitrogenadosDokumen192 halamanCompuestos NitrogenadosMilton IsraelBelum ada peringkat
- LÍPIDOSISDokumen52 halamanLÍPIDOSISED MONTBelum ada peringkat
- Ficha Clinica ImprimirrrDokumen7 halamanFicha Clinica ImprimirrrSYLVER CH QQBelum ada peringkat
- ArteroesclerosisDokumen72 halamanArteroesclerosisKatherine GallosoBelum ada peringkat
- Sistema EndocrinoDokumen5 halamanSistema EndocrinoFernando David Ayala LlorenteBelum ada peringkat
- TetradotoxinaDokumen15 halamanTetradotoxinaVíctor AlcántarBelum ada peringkat
- AA06 Estructura de Los EritrocitosDokumen55 halamanAA06 Estructura de Los EritrocitosHenry Ponce TamayBelum ada peringkat
- 3-Renov, Repara y Regener Tisu IDokumen47 halaman3-Renov, Repara y Regener Tisu IKevin PerezBelum ada peringkat
- Sangre y HemostasiaDokumen57 halamanSangre y HemostasiaMacarena Riffo WestermannBelum ada peringkat
- Bioquimica Proteinas Plasmaticas TerminadoDokumen15 halamanBioquimica Proteinas Plasmaticas Terminadoerick marioBelum ada peringkat
- Protocolo de Investigacion Relacion Entre El DX Enfermero y La Valoracio Cefalocaudal EneroDokumen35 halamanProtocolo de Investigacion Relacion Entre El DX Enfermero y La Valoracio Cefalocaudal EneroClaudia GilBelum ada peringkat
- Dislipidemia ExposicionDokumen10 halamanDislipidemia ExposicionMonica Ospina100% (1)
- 140 - Fisiología de Las Glándulas Tiroides y ParatiroidesDokumen18 halaman140 - Fisiología de Las Glándulas Tiroides y ParatiroidesAnita AlanizBelum ada peringkat
- Manual Nutricion Enfermedad Renal KelloggsDokumen22 halamanManual Nutricion Enfermedad Renal KelloggsAngie SaileBelum ada peringkat
- Células y Moléculas Del Sistema Inmune InnatoDokumen20 halamanCélulas y Moléculas Del Sistema Inmune InnatoFernando Aponte R.Belum ada peringkat
- Expo Endocrino - Hipotálamo e HipófisisDokumen93 halamanExpo Endocrino - Hipotálamo e HipófisisLupita AriasBelum ada peringkat
- Vitaminas Liposolubles e HidrosolublesDokumen19 halamanVitaminas Liposolubles e HidrosolublesAngie SaileBelum ada peringkat
- InflamaciónDokumen9 halamanInflamaciónIGNACIO SALVADOR FERNANDEZ GIRONBelum ada peringkat
- HematopoyesisDokumen13 halamanHematopoyesisLEO 999Belum ada peringkat
- DSM5 Medidas Evaluacion Entrevista Formulacion Cultural (EFC)Dokumen4 halamanDSM5 Medidas Evaluacion Entrevista Formulacion Cultural (EFC)Undomiel77Belum ada peringkat
- Dosaje de Cadenas LigerasDokumen56 halamanDosaje de Cadenas LigerasCarol TorresBelum ada peringkat
- Glándula TiroidesDokumen42 halamanGlándula TiroidesTamara Ruiz MaureiraBelum ada peringkat
- RECEPTORESDokumen5 halamanRECEPTORESMaryen Gallardo ovandoBelum ada peringkat
- Perfil Tiroideo Interpretacion PDFDokumen27 halamanPerfil Tiroideo Interpretacion PDFTalia LizardiBelum ada peringkat
- INMUNOLOGÍA - Tema 9 El TCRDokumen10 halamanINMUNOLOGÍA - Tema 9 El TCRMarcos Navarro BuiguesBelum ada peringkat
- Histo Sem 01 (Técnica Histológica)Dokumen124 halamanHisto Sem 01 (Técnica Histológica)Celso NemerBelum ada peringkat
- Deficiencia de G6P-DeshidrogenasaDokumen49 halamanDeficiencia de G6P-DeshidrogenasaCésar AmcBelum ada peringkat
- Hormonas Tiroideas y Su MetabolismoDokumen45 halamanHormonas Tiroideas y Su MetabolismoNelson PáezBelum ada peringkat
- Histología de Las Glándulas Adrenales y El PáncreasfDokumen30 halamanHistología de Las Glándulas Adrenales y El PáncreasfCamila Rubilar TroncosoBelum ada peringkat
- Sintesis, Mecanismo y Accion de La Glandula TiroideaDokumen10 halamanSintesis, Mecanismo y Accion de La Glandula Tiroideaaady fernandezBelum ada peringkat
- Membrana PlasmaticaDokumen66 halamanMembrana PlasmaticaKiara Senmache100% (1)
- Determinación de HormonasDokumen7 halamanDeterminación de HormonasSebastian RodasBelum ada peringkat
- 3212glandula Tiroidea OrtizDokumen28 halaman3212glandula Tiroidea OrtizJatnsel Reymond Alcántara CalderónBelum ada peringkat
- Resumen EndocrinoDokumen33 halamanResumen Endocrinojohana paola merlano sotoBelum ada peringkat
- Aparat YuxtaglomerularDokumen17 halamanAparat Yuxtaglomerularana paolaBelum ada peringkat
- Perfil GinecologicoDokumen18 halamanPerfil GinecologicoluzveliaBelum ada peringkat
- HipofisisDokumen18 halamanHipofisisnayeli esmeralda avalos celisBelum ada peringkat
- Glándulas Sebáceas y SudoríparasDokumen15 halamanGlándulas Sebáceas y SudoríparasMarco GonzalezBelum ada peringkat
- Citogenética y Sus AplicacionesDokumen25 halamanCitogenética y Sus AplicacionesJR Bio GenBelum ada peringkat
- HipertiroidismoDokumen34 halamanHipertiroidismoCami LaureBelum ada peringkat
- Asfixia Perinatal.Dokumen21 halamanAsfixia Perinatal.German Andres ZambranoBelum ada peringkat
- Clase 10 CitoesqueletoDokumen60 halamanClase 10 CitoesqueletoElizabethMartinezBeltranBelum ada peringkat
- Funciones Fisiolgicas de La Hormona de CrecimientoDokumen90 halamanFunciones Fisiolgicas de La Hormona de CrecimientoMauro BedoyaBelum ada peringkat
- Glandula TimoDokumen4 halamanGlandula TimoYARIBelum ada peringkat
- CitoesqueletoDokumen3 halamanCitoesqueletoAlexisBelum ada peringkat
- 2.-Tiroides ClasesDokumen28 halaman2.-Tiroides ClasesYos GOarBelum ada peringkat
- HematopoyesisDokumen15 halamanHematopoyesisHaah SgsBelum ada peringkat
- CEREBELO ResumenDokumen8 halamanCEREBELO ResumenMelina BarviBelum ada peringkat
- Perfil de LípidosDokumen31 halamanPerfil de LípidosAbril TrujilloBelum ada peringkat
- Sistema EndocrinoDokumen12 halamanSistema EndocrinoKimmi LoayzaBelum ada peringkat
- Seminario de Celulas DendriticasDokumen24 halamanSeminario de Celulas DendriticasAnonymous IuimQGqnmBelum ada peringkat
- GENERALIDADESDokumen22 halamanGENERALIDADESLizeth RíosBelum ada peringkat
- Prolactina 2Dokumen6 halamanProlactina 2Kelvin1045Belum ada peringkat
- ERRORES INNATOS DEL METABOLISMO DE LOS LDokumen42 halamanERRORES INNATOS DEL METABOLISMO DE LOS Lmarie1690Belum ada peringkat
- Receptores y Marcadores LinfocitariosDokumen16 halamanReceptores y Marcadores LinfocitariosSilvana JsbBelum ada peringkat
- Guía de Endocrino Primer Departamental 1Dokumen14 halamanGuía de Endocrino Primer Departamental 1luucy0lopez0raBelum ada peringkat
- Toma de MuestrasDokumen52 halamanToma de MuestrasShannen Mileth VizcaínoBelum ada peringkat
- Agonistas ColinérgicosDokumen13 halamanAgonistas ColinérgicosruloBelum ada peringkat
- Sindromes EndocrinosDokumen29 halamanSindromes EndocrinosAngel David Sanchez MorenoBelum ada peringkat
- Fisiopatología Del Eje Hipotálamo-Hipofisio-TiroideoDokumen22 halamanFisiopatología Del Eje Hipotálamo-Hipofisio-TiroideoMary ClaudiaBelum ada peringkat
- Fisiología de LeucocitosDokumen3 halamanFisiología de LeucocitosChristian Delgado JulcaBelum ada peringkat
- Resumen Seminario Hormonas (T3 y T4) : Natalia Klinger Chaverra. Zamira Sarmiento Chaverra. Grupo 3Dokumen5 halamanResumen Seminario Hormonas (T3 y T4) : Natalia Klinger Chaverra. Zamira Sarmiento Chaverra. Grupo 3FREDY ENRIQUE100% (1)
- Respuesta Adquirida (Inmunidad Adaptativa)Dokumen17 halamanRespuesta Adquirida (Inmunidad Adaptativa)Orbely Garcia100% (1)
- Glándula Tiroides 1Dokumen12 halamanGlándula Tiroides 1Fede AranaBelum ada peringkat
- Soporte Nutricional 2016Dokumen33 halamanSoporte Nutricional 2016Angie SaileBelum ada peringkat
- Anexos 2016Dokumen6 halamanAnexos 2016Angie SaileBelum ada peringkat
- Unidad 3 LipidosDokumen23 halamanUnidad 3 LipidosAngie SaileBelum ada peringkat
- Vitaminas y Minerales 2016Dokumen96 halamanVitaminas y Minerales 2016Angie SaileBelum ada peringkat
- Diabetes (17-08-16)Dokumen41 halamanDiabetes (17-08-16)Angie SaileBelum ada peringkat
- Unidad 2 Hidratos de Carbono y Fibra DietariaDokumen34 halamanUnidad 2 Hidratos de Carbono y Fibra DietariaAngie SaileBelum ada peringkat
- Unidad 1 ProteinasDokumen12 halamanUnidad 1 ProteinasAngie SaileBelum ada peringkat
- Sistema de Salud ArgentinoDokumen2 halamanSistema de Salud ArgentinoAngie SaileBelum ada peringkat
- Original9 - Nutricion Parenteral Pediatric ADokumen10 halamanOriginal9 - Nutricion Parenteral Pediatric ADaniela Gálvez SilvaBelum ada peringkat
- Patologias IntestinalesDokumen26 halamanPatologias IntestinalesAngie SaileBelum ada peringkat
- Clase 4 Actividad Fisica y SaludDokumen106 halamanClase 4 Actividad Fisica y SaludAngie SaileBelum ada peringkat
- Preparacion Fisica de NadadoresDokumen8 halamanPreparacion Fisica de NadadoresCarlos Manuel BorjaBelum ada peringkat
- Clase 2 EnergiaDokumen39 halamanClase 2 EnergiaAngie SaileBelum ada peringkat
- Sistema de Salud ArgentinoDokumen2 halamanSistema de Salud ArgentinoAngie SaileBelum ada peringkat
- Prueba de RombergDokumen5 halamanPrueba de RombergAngie SaileBelum ada peringkat
- Abordaje Analisis Situacion SaludDokumen31 halamanAbordaje Analisis Situacion SaludAngie SaileBelum ada peringkat
- Macronutrientes PDFDokumen102 halamanMacronutrientes PDFLis VibeBelum ada peringkat
- Plantas - Enciclopedia Del JardinDokumen351 halamanPlantas - Enciclopedia Del JardinOscar Rojas100% (1)
- 2015-11 Mortalidad Tabla-3 Msal - GobDokumen2 halaman2015-11 Mortalidad Tabla-3 Msal - GobAngie SaileBelum ada peringkat
- Guia 1Dokumen6 halamanGuia 1Angie SaileBelum ada peringkat
- Fibrosis QuísticaDokumen10 halamanFibrosis QuísticaAngie SaileBelum ada peringkat
- 2009 Receptores SensorialesDokumen5 halaman2009 Receptores SensorialesAngie SaileBelum ada peringkat
- Organización Del Sistema EndócrinoDokumen7 halamanOrganización Del Sistema EndócrinoAngie SaileBelum ada peringkat
- Fibrosis QuísticaDokumen10 halamanFibrosis QuísticaAngie SaileBelum ada peringkat
- Cuestionario 2Dokumen10 halamanCuestionario 2yuliet mendoza40% (5)
- Deshidratacion AlbayDokumen22 halamanDeshidratacion Albayconsultorio la cardeneraBelum ada peringkat
- Tumores MalignosDokumen19 halamanTumores MalignosValerie SandraBelum ada peringkat
- Clasificación DisartriaDokumen31 halamanClasificación DisartriaAlondra Villagran100% (1)
- IsocianatoDokumen1 halamanIsocianatoTatiana GuashpaBelum ada peringkat
- Contenido Programatico Infecciosas I 2020-2Dokumen14 halamanContenido Programatico Infecciosas I 2020-2Mike SanchezBelum ada peringkat
- Patologia ClinicaDokumen224 halamanPatologia Clinicamotanet100% (1)
- Documento Final InfluenzaDokumen61 halamanDocumento Final InfluenzaNathaly Velasteguí PeraltaBelum ada peringkat
- Rat 012012Dokumen70 halamanRat 012012dane0% (1)
- Manejo Sanitario Del Rebaño Caprino (S. Nieto y W Armas)Dokumen19 halamanManejo Sanitario Del Rebaño Caprino (S. Nieto y W Armas)Alirio BoscánBelum ada peringkat
- 7ma Clase - Quinolonas, Sulfonamidas, Trimetoprim. Dra. de FreitasDokumen39 halaman7ma Clase - Quinolonas, Sulfonamidas, Trimetoprim. Dra. de FreitasCátedra de Farmacología de la Escuela de Medicina de La UNIVERSIDAD DEL ZULIA100% (2)
- Mecanismos de Muerte CelularDokumen11 halamanMecanismos de Muerte CelularJANELA KATHERINE MUÑOZ ROSASBelum ada peringkat
- Las Toxinas Ambientales y Sus Efectos GenéticosDokumen78 halamanLas Toxinas Ambientales y Sus Efectos Genéticosarcadia_hb100% (1)
- Lesión Medular y Trauma Craneoncefalico Severo-1Dokumen19 halamanLesión Medular y Trauma Craneoncefalico Severo-1JUAN PABLO MORALES BASTOBelum ada peringkat
- 1025 5583 Afm 82 01 00021Dokumen6 halaman1025 5583 Afm 82 01 00021dianabrito72273928Belum ada peringkat
- Determinación de La Edad en EquinosDokumen7 halamanDeterminación de La Edad en EquinosChris OlivaresBelum ada peringkat
- Informe Farmacos para El Asma y Epoc - Grupo 4Dokumen23 halamanInforme Farmacos para El Asma y Epoc - Grupo 4Druman VélezBelum ada peringkat
- Apuntes InmunologíaDokumen21 halamanApuntes InmunologíaanonimousBelum ada peringkat
- Equipo 7 - Ev2 - Intro A La MedDokumen4 halamanEquipo 7 - Ev2 - Intro A La MeddiegodavilaaguilarBelum ada peringkat
- Evacuacion Inicial Cancer InfantilDokumen6 halamanEvacuacion Inicial Cancer InfantilJazmin TorresBelum ada peringkat
- The BandhasDokumen7 halamanThe BandhasLeo CasariniBelum ada peringkat
- CardiovascularDokumen60 halamanCardiovascularMariana RojasBelum ada peringkat
- Mecanismos de DefensaDokumen21 halamanMecanismos de Defensabrayan nicolas monroy camachoBelum ada peringkat
- Formato de Dosificación Enero 2023 BIOLOGIADokumen7 halamanFormato de Dosificación Enero 2023 BIOLOGIAEmilio PérezBelum ada peringkat
- Protocolo Aislamiento de Agentes Patógenos Medicina Interna HNHUDokumen14 halamanProtocolo Aislamiento de Agentes Patógenos Medicina Interna HNHUVictor Guerrero QuijanoBelum ada peringkat
- Pretest Rm3v Salud Publica Sin ClaveDokumen3 halamanPretest Rm3v Salud Publica Sin ClavesaidajaraBelum ada peringkat
- Resultados Vivero La Jugoza Nov17Dokumen3 halamanResultados Vivero La Jugoza Nov17Lina CamposBelum ada peringkat