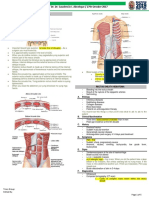
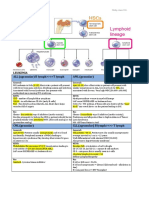
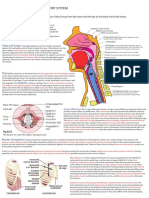
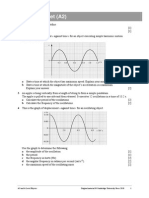
Anda mungkin juga menyukai
- Cytogenetics Basics: Chromosomes, Analysis & AbnormalitiesDokumen11 halamanCytogenetics Basics: Chromosomes, Analysis & Abnormalitiesjo_jo_mania100% (1)
- Biliary Disorders and Pancreatic Conditions ExplainedDokumen59 halamanBiliary Disorders and Pancreatic Conditions ExplainedErick Sioco InsertoBelum ada peringkat
- Genetics: OLFU - College of MedicineDokumen7 halamanGenetics: OLFU - College of MedicineMiguel CuevasBelum ada peringkat
- Pelvis and Perineum Clinical CorrelationDokumen4 halamanPelvis and Perineum Clinical CorrelationKeesha Mariel AlimonBelum ada peringkat
- Abdominal Wall, Omentum, Mesentery, Retroperitoneum Anatomy and ConditionsDokumen6 halamanAbdominal Wall, Omentum, Mesentery, Retroperitoneum Anatomy and ConditionsMon Ordona De GuzmanBelum ada peringkat
- SYPHYLISDokumen1 halamanSYPHYLISkhadzxBelum ada peringkat
- Brainstem Bravo AnnotatedDokumen13 halamanBrainstem Bravo AnnotatedMia CadizBelum ada peringkat
- Anatomy Formative Preview of Thorax and BackDokumen22 halamanAnatomy Formative Preview of Thorax and Backmvs hardhikBelum ada peringkat
- Risk Factors - Step 2 PDFDokumen5 halamanRisk Factors - Step 2 PDFAmberBelum ada peringkat
- Essential Update: FDA Approves First Test To Predict AKI in Critically Ill PatientsDokumen5 halamanEssential Update: FDA Approves First Test To Predict AKI in Critically Ill PatientsRika Ariyanti SaputriBelum ada peringkat
- NephroticDokumen8 halamanNephroticsangheetaBelum ada peringkat
- MCB 252 Final Exam Study GuideDokumen62 halamanMCB 252 Final Exam Study GuideJay ZBelum ada peringkat
- Aquifer Case - Summary - FamilyMedicine32 - 33-YDokumen9 halamanAquifer Case - Summary - FamilyMedicine32 - 33-YHyunsoo EllisBelum ada peringkat
- Glomerular DiseaseDokumen98 halamanGlomerular DiseasePatrick TumewuBelum ada peringkat
- 2011 07 Microbiology Mycobacterium Skin InfectionDokumen6 halaman2011 07 Microbiology Mycobacterium Skin InfectionCristinaConcepcionBelum ada peringkat
- Aquifer Case - Summary - FamilyMedicine21 - 12-YDokumen10 halamanAquifer Case - Summary - FamilyMedicine21 - 12-YHyunsoo EllisBelum ada peringkat
- Aquifer Case 27Dokumen4 halamanAquifer Case 27Dina KristevaBelum ada peringkat
- NSAID's "Non-Steroidal Anti-Inflammatory Drugs": Mmbakhaitan@uqu - Edu.saDokumen19 halamanNSAID's "Non-Steroidal Anti-Inflammatory Drugs": Mmbakhaitan@uqu - Edu.saAhmed HossamBelum ada peringkat
- Chronic Inflammatory Dermatoses Inflammatory Blistering DisordersDokumen4 halamanChronic Inflammatory Dermatoses Inflammatory Blistering DisordersspringdingBelum ada peringkat
- Skin Structure and Function GuideDokumen10 halamanSkin Structure and Function GuideyassrmarwaBelum ada peringkat
- Renal SyndromeDokumen13 halamanRenal SyndromeAndreas KristianBelum ada peringkat
- Aquifer Study CaseDokumen8 halamanAquifer Study CaseRosanaBelum ada peringkat
- Actinic Keratosis: (Aka Bowen's Disease)Dokumen5 halamanActinic Keratosis: (Aka Bowen's Disease)fadoBelum ada peringkat
- Nephrotic Syndrome WikipediaDokumen10 halamanNephrotic Syndrome WikipediaJohn KevlarBelum ada peringkat
- 5 Cyto AbnormalDokumen9 halaman5 Cyto AbnormalMerli Ann Joyce CalditoBelum ada peringkat
- Non-Pathogenic Intestinal Amoebae Cyst MorphologyDokumen2 halamanNon-Pathogenic Intestinal Amoebae Cyst MorphologyCoy NuñezBelum ada peringkat
- Embryology - PharyngealDokumen11 halamanEmbryology - PharyngealKhoa Thanh Nguyen Le100% (1)
- Abdominal Pain ExplainedDokumen4 halamanAbdominal Pain ExplainedNazmun LailahBelum ada peringkat
- Nephrotic Nephritic SyndromsDokumen4 halamanNephrotic Nephritic SyndromsKimiwari100% (2)
- Histo Review 2Dokumen13 halamanHisto Review 2Coy NuñezBelum ada peringkat
- Cycle CellDokumen16 halamanCycle CellRohingya EnglishBelum ada peringkat
- Goljan Audios Transcribed (Margaret)Dokumen216 halamanGoljan Audios Transcribed (Margaret)sean8phamBelum ada peringkat
- Upper and Lower Limb NervesDokumen6 halamanUpper and Lower Limb NervesTaimoor DogarBelum ada peringkat
- Anterior Abdominal Wall Surgery InsightsDokumen5 halamanAnterior Abdominal Wall Surgery InsightsAnonymous 2TzM1ZBelum ada peringkat
- Head & Neck - Deep Cervical Fascia &cervical PlexusDokumen9 halamanHead & Neck - Deep Cervical Fascia &cervical PlexusKomang Tri Adi SuparwatiBelum ada peringkat
- Inguinal CanalDokumen4 halamanInguinal CanalspiraldaoBelum ada peringkat
- Pathology Finals Reviewer on WBCs, Lymph Nodes, Spleen & ThymusDokumen5 halamanPathology Finals Reviewer on WBCs, Lymph Nodes, Spleen & Thymusangel_sagun_1Belum ada peringkat
- Patho CA - Acute PancreatitisDokumen1 halamanPatho CA - Acute PancreatitisKBelum ada peringkat
- Pathology B - Gastrointestinal Tract (Esguerra, 2015)Dokumen18 halamanPathology B - Gastrointestinal Tract (Esguerra, 2015)Ars MoriendiBelum ada peringkat
- Heart Failure - Notes From "Cardiology" (Timmis Et Al) : Main CausesDokumen3 halamanHeart Failure - Notes From "Cardiology" (Timmis Et Al) : Main CausesPrarthana Thiagarajan100% (3)
- Leukemia and Lymphoma OverviewDokumen2 halamanLeukemia and Lymphoma OverviewAyeshaArifBelum ada peringkat
- Kidney Physiology (Q & A)Dokumen28 halamanKidney Physiology (Q & A)ramadan100% (1)
- VirologyLearnTable 2017-Sketchy PDFDokumen4 halamanVirologyLearnTable 2017-Sketchy PDFdfgbdfgBelum ada peringkat
- Introduction To TransplantationDokumen3 halamanIntroduction To TransplantationGerardLumBelum ada peringkat
- Identify viscus from histological featuresDokumen1 halamanIdentify viscus from histological featuresNaser Hamdi ZalloumBelum ada peringkat
- Respiratory Notes (Chris Andersen, ICUPrimaryPrep - Com)Dokumen14 halamanRespiratory Notes (Chris Andersen, ICUPrimaryPrep - Com)PkernBelum ada peringkat
- Disease PDFDokumen6 halamanDisease PDFJohn Christopher LucesBelum ada peringkat
- Genetic Disorders-Www - Qworld.co - inDokumen13 halamanGenetic Disorders-Www - Qworld.co - inQworld100% (1)
- Divine Intervention Episode 12 Bacteria Cases ADokumen18 halamanDivine Intervention Episode 12 Bacteria Cases ASwisskelly1Belum ada peringkat
- General Features of The Immune SystemDokumen6 halamanGeneral Features of The Immune SystemMinerva Bautista RoseteBelum ada peringkat
- 1 Ana Intro Finals September 16 LaygoDokumen3 halaman1 Ana Intro Finals September 16 LaygombdelenaBelum ada peringkat
- The Immune System DefencesDokumen6 halamanThe Immune System DefencesKa-Shun Leung100% (1)
- Immune Disorders ExplainedDokumen5 halamanImmune Disorders ExplainedThalia Fortune100% (1)
- Chapter 3 Genetic VariationDokumen21 halamanChapter 3 Genetic VariationAbdulkarimBelum ada peringkat
- Kidney NewDokumen4 halamanKidney NewParth BhayanaBelum ada peringkat
- 4.1d - Pathology of The Pituitary - Nov.10 - Dr. GalangDokumen4 halaman4.1d - Pathology of The Pituitary - Nov.10 - Dr. GalangMiel Raphael AranillaBelum ada peringkat
- Chromosomal AbrerationsDokumen34 halamanChromosomal AbrerationsSamar MagdyBelum ada peringkat
- Chromosome Structural ChangesDokumen51 halamanChromosome Structural ChangesdrhydrogenBelum ada peringkat
- Chromosome Inversions: Saint Mary's HospitalDokumen12 halamanChromosome Inversions: Saint Mary's HospitalMerjema BahtanovićBelum ada peringkat
- Chromosomal Abnormalities Causes and TypesDokumen14 halamanChromosomal Abnormalities Causes and TypesMerjema BahtanovićBelum ada peringkat
- Chromosomal Syndromes and Genetic DiseaseDokumen12 halamanChromosomal Syndromes and Genetic DiseasePIgnacio Alarcon AriasBelum ada peringkat
- Study-Skills-Tips DmuDokumen3 halamanStudy-Skills-Tips Dmuapi-213648940Belum ada peringkat
- Inherited Human Diseases Caused by Defect in DNA RepairDokumen1 halamanInherited Human Diseases Caused by Defect in DNA RepairMerjema BahtanovićBelum ada peringkat
- Robertsonian Chromosome Translocations ExplainedDokumen8 halamanRobertsonian Chromosome Translocations ExplainedMerjema BahtanovićBelum ada peringkat
- Robertsonian Translocations FTNWDokumen8 halamanRobertsonian Translocations FTNWRobert ChristevenBelum ada peringkat
- Chromosomal Syndromes and Genetic DiseaseDokumen12 halamanChromosomal Syndromes and Genetic DiseasePIgnacio Alarcon AriasBelum ada peringkat
- Cell Structures in DetailDokumen6 halamanCell Structures in DetailCleo PoulosBelum ada peringkat
- Gateway B2+ Student's Book Wordlist - Bosnian: Unit 1Dokumen180 halamanGateway B2+ Student's Book Wordlist - Bosnian: Unit 1Merjema Bahtanović100% (1)
- HGSS Chapter08 NewDokumen19 halamanHGSS Chapter08 NewYeti MaretaBelum ada peringkat
- 9701 Chemistry Applications BookletDokumen161 halaman9701 Chemistry Applications BookletZain RehanBelum ada peringkat
- Worksheet 20 PDFDokumen3 halamanWorksheet 20 PDFVijay BhaskarBelum ada peringkat
- 2.5 NotesDokumen10 halaman2.5 NotesxstrawberryswingxBelum ada peringkat
- Aqa As Chemsitry Chapter 3Dokumen29 halamanAqa As Chemsitry Chapter 3khan.inayat2Belum ada peringkat
- Tackling ExamDokumen17 halamanTackling ExamIbad MohammedBelum ada peringkat
- Physics Unit 2 NotesDokumen3 halamanPhysics Unit 2 NotesSmallville96100% (1)
- LymphocyteDokumen5 halamanLymphocyteMerjema BahtanovićBelum ada peringkat
- 9700 s13 QP 23Dokumen12 halaman9700 s13 QP 23KimberlyLaw95Belum ada peringkat
- 9700 s13 QP 23Dokumen12 halaman9700 s13 QP 23KimberlyLaw95Belum ada peringkat
- 9701 Chemistry Applications BookletDokumen161 halaman9701 Chemistry Applications BookletZain RehanBelum ada peringkat
- Bio 201 Chapter Objectives For Chapters 22 - 25Dokumen17 halamanBio 201 Chapter Objectives For Chapters 22 - 25Thomas Trotter100% (6)
- Anthurium Breeding PPT by S Y ChandrashekarDokumen30 halamanAnthurium Breeding PPT by S Y ChandrashekarDr-Chandrashekar S Y100% (20)
- Biochem Genetics USMLE Practice Questions (2 of 4)Dokumen13 halamanBiochem Genetics USMLE Practice Questions (2 of 4)MedSchoolStuff100% (2)
- Hiphedera ALDokumen154 halamanHiphedera ALMichelle DulayBelum ada peringkat
- Biology Notes Form 4 PDF - Biology Form 4 Questions and Answers PDFDokumen94 halamanBiology Notes Form 4 PDF - Biology Form 4 Questions and Answers PDFjalaldinmohammed201Belum ada peringkat
- 8-Abnormal-Chromosome-Number 2Dokumen34 halaman8-Abnormal-Chromosome-Number 2PCHS-REYES, Reyza Jane BBelum ada peringkat
- GOTA Colchicine - Wikipedia, The Free EncyclopediaDokumen9 halamanGOTA Colchicine - Wikipedia, The Free Encyclopediavaleriano pedroBelum ada peringkat
- Introduction To Biotechnology 3Rd Edition Thieman Test Bank Full Chapter PDFDokumen12 halamanIntroduction To Biotechnology 3Rd Edition Thieman Test Bank Full Chapter PDFchastescurf7btc100% (7)
- 10 11648 J Ajpb S 2017020301 13Dokumen5 halaman10 11648 J Ajpb S 2017020301 13mufida chairunisaBelum ada peringkat
- Protoplast CultureDokumen45 halamanProtoplast CultureRabia Ahmed100% (1)
- Published PaperDokumen11 halamanPublished PaperManjit KaurBelum ada peringkat
- Biology Chapter 15Dokumen15 halamanBiology Chapter 15huifwiu jsbBelum ada peringkat
- Species Concepts ExplainedDokumen26 halamanSpecies Concepts ExplainedmashedtomatoezedBelum ada peringkat
- Chapter 17 - Griffiths An Introduction To Genetic Analysis 11th Edition 2015Dokumen47 halamanChapter 17 - Griffiths An Introduction To Genetic Analysis 11th Edition 2015Xime RdzBelum ada peringkat
- Understanding Pathophysiology 6th Edition Huether Test BankDokumen11 halamanUnderstanding Pathophysiology 6th Edition Huether Test Bankheatherowensrtpcewjmag100% (16)
- Tife Chapter Two-2Dokumen37 halamanTife Chapter Two-2Ajegbomogun SundayBelum ada peringkat
- 0 Chapter 24 Origin of Evolution Reading Guide AnswersDokumen4 halaman0 Chapter 24 Origin of Evolution Reading Guide AnswerskkkkccBelum ada peringkat
- Principles of Crop Production - Introduction LectureDokumen44 halamanPrinciples of Crop Production - Introduction LectureMarc Ice creamBelum ada peringkat
- Selection and Speciation NotesDokumen7 halamanSelection and Speciation NotesLIM ZHI SHUENBelum ada peringkat
- Chp. 24, Origin of Species1Dokumen86 halamanChp. 24, Origin of Species1zoology qauBelum ada peringkat
- A2as Biol Revised Support 2044Dokumen123 halamanA2as Biol Revised Support 2044Meagan MahangooBelum ada peringkat
- GPB 321 Crop Improvement Manual FinalDokumen71 halamanGPB 321 Crop Improvement Manual FinalSultan AhmadBelum ada peringkat
- Proceeding ISTH 2016 - 3 Juli 2017 1Dokumen176 halamanProceeding ISTH 2016 - 3 Juli 2017 1Alvian putraparlinBelum ada peringkat
- PolyploidyDokumen7 halamanPolyploidyleticiabilboBelum ada peringkat
- Chromosome VariationsDokumen39 halamanChromosome VariationsXwag 12Belum ada peringkat
- Recent Technological Innovations in Aquaculture: January 2003Dokumen17 halamanRecent Technological Innovations in Aquaculture: January 2003Jinnie R. MamhotBelum ada peringkat
- Plant Taxonomy Tools and Evidence from Palynology, Cytology, Phytochemistry and Molecular DataDokumen17 halamanPlant Taxonomy Tools and Evidence from Palynology, Cytology, Phytochemistry and Molecular DataYanie Ummu ArsylaBelum ada peringkat
- SPECIATION For FinalDokumen20 halamanSPECIATION For FinalSitti Naisa ArahamBelum ada peringkat
- BIOL2162 Course Outline March 2012Dokumen19 halamanBIOL2162 Course Outline March 2012slackerzzBelum ada peringkat
- Study Unit 5: Evolutionary Genetics: 26.1 Organisms Evolve Through Genetic Change Taking Place Within PopulationsDokumen10 halamanStudy Unit 5: Evolutionary Genetics: 26.1 Organisms Evolve Through Genetic Change Taking Place Within PopulationsKayleigh MastersBelum ada peringkat