Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Revista Ces Publicacion 2016Dokumen121 halamanRevista Ces Publicacion 2016Yenny RodriguezBelum ada peringkat
- Art07 Mathias PDFDokumen10 halamanArt07 Mathias PDFsebastianBelum ada peringkat
- Functionality of Milk Powder and Milk Based Powder For End Use Applications A ReviewDokumen11 halamanFunctionality of Milk Powder and Milk Based Powder For End Use Applications A ReviewYenny RodriguezBelum ada peringkat
- Fao 2016 The State of Food and Agriculture PDFDokumen194 halamanFao 2016 The State of Food and Agriculture PDFYenny Rodriguez100% (1)
- Aoac Methods 1 1990Dokumen771 halamanAoac Methods 1 1990Erika Lopez ArandaBelum ada peringkat
- CienciaDokumen10 halamanCienciaYenny RodriguezBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Electrochemical Cell LabDokumen9 halamanElectrochemical Cell Labribots0% (1)
- Stainless Steel Putty (ST) : Technical Data SheetDokumen2 halamanStainless Steel Putty (ST) : Technical Data SheetSreenivasBelum ada peringkat
- Flexural Testing ASTM D790Dokumen11 halamanFlexural Testing ASTM D790habiba jamilBelum ada peringkat
- Alkynes: An Introduction To Organic Synthesis: Based On Mcmurry'S Organic Chemistry, 7 EditionDokumen26 halamanAlkynes: An Introduction To Organic Synthesis: Based On Mcmurry'S Organic Chemistry, 7 EditionElisha NiñaBelum ada peringkat
- ECSS Q ST 70 15C (1may2021)Dokumen124 halamanECSS Q ST 70 15C (1may2021)Navamani Prakash100% (1)
- Ecotoxicology and Environmental SafetyDokumen10 halamanEcotoxicology and Environmental SafetyUlfa Mudia SariBelum ada peringkat
- Physico-Chemical Properties of Drinking WaterDokumen12 halamanPhysico-Chemical Properties of Drinking WaterEmmanuel WinfulBelum ada peringkat
- Fingermark Visualisation Manual Presentation IAI Aug 2013Dokumen42 halamanFingermark Visualisation Manual Presentation IAI Aug 2013Dogaru MugurBelum ada peringkat
- Sample Problem 1: X Direction X Direction XDokumen4 halamanSample Problem 1: X Direction X Direction XHarold MangaBelum ada peringkat
- Growth and Characterization of L-Alanine Potassium Nitrate Single Crystals For Nonlinear Optical ApplicationsDokumen5 halamanGrowth and Characterization of L-Alanine Potassium Nitrate Single Crystals For Nonlinear Optical ApplicationsPalaniswamy SankariahBelum ada peringkat
- Basic Laboratory Glassware and EquipmentDokumen11 halamanBasic Laboratory Glassware and EquipmentSheila Mae BaranganBelum ada peringkat
- DPP 9Dokumen3 halamanDPP 9Sarvesh DubeyBelum ada peringkat
- A Brief Overview of Fly Ash Brick Production: January 2012Dokumen13 halamanA Brief Overview of Fly Ash Brick Production: January 2012Hadi Iz'aanBelum ada peringkat
- LalruDokumen98 halamanLalruAbhinav KumarBelum ada peringkat
- Aldehyde and Ketones Tollen TestDokumen9 halamanAldehyde and Ketones Tollen TestGule RashBelum ada peringkat
- Nanoformulation of Curcuma Longa Root Extract and Evaluation of Its Dissolution PotentialDokumen9 halamanNanoformulation of Curcuma Longa Root Extract and Evaluation of Its Dissolution Potentialmuhammad adnan ayubBelum ada peringkat
- Densification of Biomass by Briquetting: A Review: October 2017Dokumen9 halamanDensification of Biomass by Briquetting: A Review: October 2017Mauri RangelBelum ada peringkat
- TWI Current Practice For Welding P92 PDFDokumen13 halamanTWI Current Practice For Welding P92 PDFpipedown456100% (1)
- Chemistry Project - A Comparison of The Solubilities of Carbon Dioxide in Water at Various TemperaturesDokumen4 halamanChemistry Project - A Comparison of The Solubilities of Carbon Dioxide in Water at Various Temperatureszaid mansuri100% (3)
- Eudragit l100 55 PDFDokumen2 halamanEudragit l100 55 PDFJohnBelum ada peringkat
- Tutorial ERP StudentsDokumen2 halamanTutorial ERP StudentsHariz MiorBelum ada peringkat
- All Aluminium Alloy ConductorsDokumen2 halamanAll Aluminium Alloy ConductorsNeeraj AgarwalBelum ada peringkat
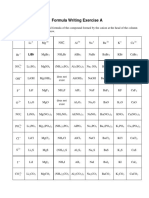
- Formula Writing - CambridgeDokumen5 halamanFormula Writing - CambridgeQusai Saify100% (3)
- Emerging Microreaction Systems Based On 3D PrintinDokumen11 halamanEmerging Microreaction Systems Based On 3D PrintinMaria Cristina López AreizaBelum ada peringkat
- Azbil LX7000 Series (Switch) - M C 2Dokumen12 halamanAzbil LX7000 Series (Switch) - M C 2phuoc leBelum ada peringkat
- NT Mat 003 - Assessment of Corrosion Protection Classes For Inorganic Coatings On Steel - Nordtest MethodDokumen7 halamanNT Mat 003 - Assessment of Corrosion Protection Classes For Inorganic Coatings On Steel - Nordtest MethodCorina StanculescuBelum ada peringkat
- Metabolism Exam 2 - GIFT - Spring 2016Dokumen9 halamanMetabolism Exam 2 - GIFT - Spring 2016shafa_nathani100% (2)
- Physical Chemistry - R. L. MadanDokumen1 halamanPhysical Chemistry - R. L. MadanOscar Santos EstofaneroBelum ada peringkat
- Introduction To Metal EtchingDokumen2 halamanIntroduction To Metal EtchingfoobiedotBelum ada peringkat
- 30 Sewage Treatment PlanDokumen63 halaman30 Sewage Treatment PlanYuri Duri100% (1)