Anda mungkin juga menyukai
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsDari EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneBelum ada peringkat
- Cell SignallingDokumen72 halamanCell SignallingSheerin SulthanaBelum ada peringkat
- Medicinal Chemistry—III: Main Lectures Presented at the Third International Symposium on Medicinal ChemistryDari EverandMedicinal Chemistry—III: Main Lectures Presented at the Third International Symposium on Medicinal ChemistryP. PratesiBelum ada peringkat
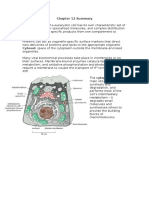
- Chapter 18 SummaryDokumen7 halamanChapter 18 SummaryCharlotteBelum ada peringkat
- Lowe Syndrome (Oculocerebrorenal syndrome) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsDari EverandLowe Syndrome (Oculocerebrorenal syndrome) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsBelum ada peringkat
- Chapter 4 SummaryDokumen8 halamanChapter 4 SummaryCharlotteBelum ada peringkat
- The Plasma Proteins V5: Structure, Function, and Genetic ControlDari EverandThe Plasma Proteins V5: Structure, Function, and Genetic ControlFrank PutnamPenilaian: 3 dari 5 bintang3/5 (1)
- Chapter 16 SummaryDokumen11 halamanChapter 16 SummaryCharlotteBelum ada peringkat
- Chapter 20 Solutions Molecular Biology of The CellDokumen15 halamanChapter 20 Solutions Molecular Biology of The CellAkshey BadrinarayanBelum ada peringkat
- Biochemistry FinalsDokumen11 halamanBiochemistry FinalsKc CalderonBelum ada peringkat
- Genome & Personality-2ndDokumen28 halamanGenome & Personality-2ndVrendra Kumar SharmaBelum ada peringkat
- Basic Molecular Biology & Biotechnology Training ManualDokumen23 halamanBasic Molecular Biology & Biotechnology Training Manualsameer420Belum ada peringkat
- Chapter 6 SummaryDokumen28 halamanChapter 6 SummaryCharlotteBelum ada peringkat
- Epidemiology 2 PDFDokumen11 halamanEpidemiology 2 PDFJawad TatlahBelum ada peringkat
- Chapter 13 SummaryDokumen5 halamanChapter 13 SummaryCharlotteBelum ada peringkat
- Chapter 12 SummaryDokumen7 halamanChapter 12 SummaryCharlotteBelum ada peringkat
- Amino Acid: Prepared By: Anthonette Ann Alemania-STEM 12Dokumen28 halamanAmino Acid: Prepared By: Anthonette Ann Alemania-STEM 12Anthonette Alemania100% (1)
- BASC02 Full PDFDokumen70 halamanBASC02 Full PDFndarubagusBelum ada peringkat
- Enzyme Inhibitors Explained: Reversible vs IrreversibleDokumen5 halamanEnzyme Inhibitors Explained: Reversible vs IrreversiblebelabelawBelum ada peringkat
- Signal Transduction Mechanisms and Cellular ResponsesDokumen39 halamanSignal Transduction Mechanisms and Cellular ResponsesBellony Sanders100% (1)
- Final Exam Study Guide TipsDokumen22 halamanFinal Exam Study Guide Tipsrazgriz1211Belum ada peringkat
- Unit 1: Introduction: Organic InorganicDokumen1 halamanUnit 1: Introduction: Organic InorganicEllah GutierrezBelum ada peringkat
- Human Genetics: From DNA to ProteinsDokumen28 halamanHuman Genetics: From DNA to ProteinsjasBelum ada peringkat
- Test 4 Essay Questoin Meiosis and GametogenesisDokumen2 halamanTest 4 Essay Questoin Meiosis and GametogenesisInactiveAccount80% (5)
- Mammalian Histology AssignmentDokumen9 halamanMammalian Histology AssignmentSana Sultana100% (1)
- Histology Notes 2Dokumen13 halamanHistology Notes 2Lukas BüchlerBelum ada peringkat
- Waves As PhysicsDokumen8 halamanWaves As PhysicsRija MughalBelum ada peringkat
- CH 10 Gene ExpressionDokumen23 halamanCH 10 Gene ExpressionerichaasBelum ada peringkat
- Cell Signaling IDokumen21 halamanCell Signaling IGuhanBelum ada peringkat
- Multidimensional Analysis of Clinical Symptoms in Patients With Fabry's DiseaseDokumen12 halamanMultidimensional Analysis of Clinical Symptoms in Patients With Fabry's DiseaseOnon EssayedBelum ada peringkat
- CNS Physiology LectureDokumen25 halamanCNS Physiology Lectureجبار هاوش عضيرس100% (1)
- Biochemistry Notes For BoardsDokumen35 halamanBiochemistry Notes For BoardsUjjwal PyakurelBelum ada peringkat
- Cell Signalling 1.2 2017Dokumen61 halamanCell Signalling 1.2 2017PutriBelum ada peringkat
- Ilovepdf MergedDokumen119 halamanIlovepdf Mergedkushal100% (1)
- EnzymesDokumen7 halamanEnzymesYoni Lacanilao100% (1)
- 4.03 Intracellular Traffic, Protein Sorting and BiosignalingDokumen16 halaman4.03 Intracellular Traffic, Protein Sorting and BiosignalingMiguel DomingoBelum ada peringkat
- 11 Immunology - ATFDokumen35 halaman11 Immunology - ATFLucas BarbosaBelum ada peringkat
- Intracellular SignallingDokumen45 halamanIntracellular Signallingsana iqbalBelum ada peringkat
- Cell Biology Review: Blood Smear and Connective TissueDokumen15 halamanCell Biology Review: Blood Smear and Connective TissueGabriel ParasBelum ada peringkat
- Examn of Molecular BiologyDokumen11 halamanExamn of Molecular BiologycegfracBelum ada peringkat
- Cell JunctionsDokumen56 halamanCell JunctionsDrGagandeep Singh100% (3)
- Neural Tube DefectsDokumen12 halamanNeural Tube Defectsdaniel_1592Belum ada peringkat
- Cell Adhesion MoleculesDokumen14 halamanCell Adhesion MoleculesSecret Agent100% (1)
- DNA Structure and Replication ExplainedDokumen12 halamanDNA Structure and Replication ExplainedJhune Dominique GalangBelum ada peringkat
- Mechanisms of Hormone Action NotesDokumen3 halamanMechanisms of Hormone Action Notesapi-390361165Belum ada peringkat
- Ch11 - Cell Signaling SlidesDokumen36 halamanCh11 - Cell Signaling SlidesValine Cysteine MethionineBelum ada peringkat
- Genetic Control of Cell Function and InheritanceDokumen7 halamanGenetic Control of Cell Function and InheritanceSADAVEJO100% (1)
- Cell Cycle Cell Division PDFDokumen13 halamanCell Cycle Cell Division PDFmuzamil shabir100% (1)
- 4 - Cell JunctionsDokumen25 halaman4 - Cell JunctionsKhdhir M. Salem100% (1)
- Characteristics and Genotyping (Semi-Automated and Automated), Apparatus Used in GenotypingDokumen45 halamanCharacteristics and Genotyping (Semi-Automated and Automated), Apparatus Used in GenotypingKhalid HameedBelum ada peringkat
- Cell SignallingDokumen37 halamanCell SignallingSreekarWunnava100% (1)
- Regulation of the Cell Cycle by Protein KinasesDokumen5 halamanRegulation of the Cell Cycle by Protein KinasesmohsinBelum ada peringkat
- Chapter 19 LodishDokumen52 halamanChapter 19 LodishablucBelum ada peringkat
- Mitosis Meiosis NOTESDokumen11 halamanMitosis Meiosis NOTESCaden DiazBelum ada peringkat
- Cellular Molecular BiologyDokumen4 halamanCellular Molecular BiologyCpopBelum ada peringkat
- MC Chapter 48 TestDokumen14 halamanMC Chapter 48 Testbori0905100% (2)
- DNA: The Genetic Material: A Molecular Approach 2 EditionDokumen63 halamanDNA: The Genetic Material: A Molecular Approach 2 Editionhomerina933379Belum ada peringkat
- Dna Genes Chromosomes 2011Dokumen65 halamanDna Genes Chromosomes 2011Suliman GarallehBelum ada peringkat
- Chromatin RemodelingDokumen5 halamanChromatin RemodelingRohit GargBelum ada peringkat
- Cell To Cell CommunicationDokumen23 halamanCell To Cell CommunicationKunal KatyayanBelum ada peringkat
- Chapter 20 SummaryyDokumen21 halamanChapter 20 SummaryyCharlotteBelum ada peringkat
- Chapter 16 SummaryDokumen11 halamanChapter 16 SummaryCharlotteBelum ada peringkat
- Chapter 19 SummaryDokumen10 halamanChapter 19 SummaryCharlotteBelum ada peringkat
- Chapter 17 SummaryDokumen22 halamanChapter 17 SummaryCharlotteBelum ada peringkat
- Chapter 3 SummaryDokumen10 halamanChapter 3 SummaryCharlotteBelum ada peringkat
- Chapter 13 SummaryDokumen5 halamanChapter 13 SummaryCharlotteBelum ada peringkat
- Chapter 8 SummaryDokumen25 halamanChapter 8 SummaryCharlotteBelum ada peringkat
- Chapter 7 SummaryDokumen34 halamanChapter 7 SummaryCharlotteBelum ada peringkat
- Chapter 12 SummaryDokumen7 halamanChapter 12 SummaryCharlotteBelum ada peringkat
- Chapter 2 SummaryDokumen3 halamanChapter 2 SummaryCharlotteBelum ada peringkat
- Chapter 6 SummaryDokumen28 halamanChapter 6 SummaryCharlotteBelum ada peringkat
- Chapter 1 SummaryDokumen4 halamanChapter 1 SummaryCharlotteBelum ada peringkat
- Crystal Appearance/Characteristics Clinical SignificanceDokumen4 halamanCrystal Appearance/Characteristics Clinical SignificanceAngela LaglivaBelum ada peringkat
- Quiz-Alcohol Ether & Phenols-Rsk - RGVDokumen6 halamanQuiz-Alcohol Ether & Phenols-Rsk - RGVAtharva GanjuBelum ada peringkat
- TRC - Lesson Plan Plant and Animal Cell 2Dokumen10 halamanTRC - Lesson Plan Plant and Animal Cell 2Armaan0% (1)
- Marble and Granite Study ReportDokumen15 halamanMarble and Granite Study Reportp156426 Ahmad KhanzadaBelum ada peringkat
- 8.2 (111 Marks) : MarkschemeDokumen47 halaman8.2 (111 Marks) : MarkschemeSemwezi Enock0% (1)
- Multistage Boiler Feed Water PumpDokumen12 halamanMultistage Boiler Feed Water Pumpbalaji_jayadeva9546100% (1)
- 4 1 - 16 10 Lws Preventers Installation and OperationDokumen28 halaman4 1 - 16 10 Lws Preventers Installation and Operationcrni roko100% (1)
- Repair and Rehabilitation of Reinforced ConcreteDokumen15 halamanRepair and Rehabilitation of Reinforced ConcreteebinVettuchirayil100% (2)
- Treating Sullage Water Using Natural and Chemical CoagulantsDokumen5 halamanTreating Sullage Water Using Natural and Chemical CoagulantsMuhammad Ramadhan SaputraBelum ada peringkat
- रासायनिक सूत्र (Chemical Formula) - Padhobeta.com Blog PDFDokumen3 halamanरासायनिक सूत्र (Chemical Formula) - Padhobeta.com Blog PDFRAKISHO WORLDBelum ada peringkat
- Id35 Honda GX630 GX660 GX690Dokumen20 halamanId35 Honda GX630 GX660 GX690RodrigoChaves0% (1)
- Disinfect water with UV lightDokumen16 halamanDisinfect water with UV lightsleonBelum ada peringkat
- Pressure Operated Water Valve Types WVFX and WVS: Data SheetDokumen11 halamanPressure Operated Water Valve Types WVFX and WVS: Data SheetДаниіл ЩигельськийBelum ada peringkat
- Product Specification: Leminar Air-Conditioning Industries LLCDokumen8 halamanProduct Specification: Leminar Air-Conditioning Industries LLCMaher Al LobnanyBelum ada peringkat
- Technical Bulletin: Ammonium Sulfate Solution, 4.1 MDokumen4 halamanTechnical Bulletin: Ammonium Sulfate Solution, 4.1 MToe PaingBelum ada peringkat
- Crack Control for Concrete Masonry VeneersDokumen10 halamanCrack Control for Concrete Masonry VeneersAdam JonesBelum ada peringkat
- Spring Steel 1Dokumen17 halamanSpring Steel 1Manicharan100% (1)
- FE Thermodynamics 7th Appendices: Water Property TablesDokumen71 halamanFE Thermodynamics 7th Appendices: Water Property TableshollegansBelum ada peringkat
- Derouging & Passivation Webinar - Q&A PDFDokumen4 halamanDerouging & Passivation Webinar - Q&A PDFDheeraj Chowdary DhanekulaBelum ada peringkat
- Jak1 Aur El DWG A 0 9035 (D)Dokumen1 halamanJak1 Aur El DWG A 0 9035 (D)Budi HahnBelum ada peringkat
- Understanding Mass SpectrometryDokumen147 halamanUnderstanding Mass SpectrometryYee Kin WengBelum ada peringkat
- Herbs MedicineDokumen237 halamanHerbs MedicineSih Kir Nugroho100% (13)
- ChemistryDokumen188 halamanChemistrySamveg ClassesBelum ada peringkat
- Piht0f402 Icmts Icad600tsDokumen2 halamanPiht0f402 Icmts Icad600tsamruta21Belum ada peringkat
- ASTM D7777 - Densimetro Digital PortatilDokumen7 halamanASTM D7777 - Densimetro Digital PortatilAlvaro Correa100% (2)
- Material Data Sheet: Urban@plastum - CZ WWW - Plastum.czDokumen1 halamanMaterial Data Sheet: Urban@plastum - CZ WWW - Plastum.czDavis GBelum ada peringkat
- Wang and Helliwell (2001) Determination of Flavonols in Green and Black Tea LeavesDokumen5 halamanWang and Helliwell (2001) Determination of Flavonols in Green and Black Tea LeavesYvonne TongBelum ada peringkat
- OTC 15275 Design Challenges of A 48-Inch Pipeline Subsea ManifoldDokumen7 halamanOTC 15275 Design Challenges of A 48-Inch Pipeline Subsea Manifoldgvsha88Belum ada peringkat
- Iccesd 2020 PDFDokumen12 halamanIccesd 2020 PDFzeekoBelum ada peringkat
- The Effect of Different Fertilizers On Plant GrowthDokumen2 halamanThe Effect of Different Fertilizers On Plant GrowthVeerath தமிழன்0% (2)