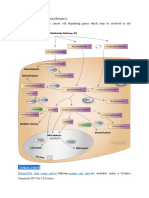
Anda mungkin juga menyukai
- JMC2013 DFG Kinase InhibitorsDokumen15 halamanJMC2013 DFG Kinase InhibitorsVincent GeruszBelum ada peringkat
- Cancers 11 00437 v3Dokumen20 halamanCancers 11 00437 v3[161]Shuaib AktherBelum ada peringkat
- Biomedicine & PharmacotherapyDokumen18 halamanBiomedicine & PharmacotherapyMiriam Sanchez moralesBelum ada peringkat
- Bioorganic & Medicinal Chemistry Letters: A 1 A 1 A B B A ADokumen9 halamanBioorganic & Medicinal Chemistry Letters: A 1 A 1 A B B A AWalid EbaiedBelum ada peringkat
- KongDokumen6 halamanKongapi-390257815Belum ada peringkat
- Kaito Et Al. - 2021 - CD155 and CD112 As Possible Therapeutic Targets ofDokumen12 halamanKaito Et Al. - 2021 - CD155 and CD112 As Possible Therapeutic Targets ofpablopac2021Belum ada peringkat
- Addition To MusculoskeletalDokumen2 halamanAddition To MusculoskeletalDrSingh1699Belum ada peringkat
- Bioorganic & Medicinal Chemistry LettersDokumen5 halamanBioorganic & Medicinal Chemistry Lettersneha bhatiaBelum ada peringkat
- RsicDokumen11 halamanRsicannounasma7Belum ada peringkat
- Bcr-Abl Tyrosine-Kinase InhibitorDokumen17 halamanBcr-Abl Tyrosine-Kinase Inhibitorolivia523Belum ada peringkat
- Ifosfamide PathwayDokumen6 halamanIfosfamide PathwayDiana AhmadBelum ada peringkat
- Sciencedirect: ArticleinfoDokumen7 halamanSciencedirect: ArticleinfoSergeat18BBelum ada peringkat
- tmp3AAA TMPDokumen10 halamantmp3AAA TMPFrontiersBelum ada peringkat
- 2019 Identifying Novel Targets of Oncogenic EGF Receptor Signaling in Lung Cancer Through Global PhosphoproteomicsDokumen16 halaman2019 Identifying Novel Targets of Oncogenic EGF Receptor Signaling in Lung Cancer Through Global Phosphoproteomicsmorteza hosseiniBelum ada peringkat
- Mol Cancer Ther-2011-Zhao-2415-25Dokumen12 halamanMol Cancer Ther-2011-Zhao-2415-25Rahul B SBelum ada peringkat
- Crystal Structures of The Kinase Domain of C-Abl in Complex With The Small Molecule Inhibitors Pd173955 and ImatinibDokumen9 halamanCrystal Structures of The Kinase Domain of C-Abl in Complex With The Small Molecule Inhibitors Pd173955 and ImatinibMarieBelum ada peringkat
- 2012AUTO0034R3Dokumen20 halaman2012AUTO0034R3rosulusBelum ada peringkat
- Synthesis and Biological Evaluation of Stilbene-Based Pure Estrogen AntagonistsDokumen5 halamanSynthesis and Biological Evaluation of Stilbene-Based Pure Estrogen AntagonistsRetno Rela MahananiBelum ada peringkat
- Inhibidores FLT3Dokumen18 halamanInhibidores FLT3Residencia PediatriaBelum ada peringkat
- Tugas Bu AniDokumen23 halamanTugas Bu AniYon-SyuhandaBelum ada peringkat
- Singh 2016Dokumen12 halamanSingh 2016Tharuka ChandrasiriBelum ada peringkat
- 04 Full Bharti MD s11030-023-10608-8Dokumen21 halaman04 Full Bharti MD s11030-023-10608-8bharti.srf.phe22Belum ada peringkat
- 2 Future Medicinal ChemistryDokumen18 halaman2 Future Medicinal ChemistryPooja NaikBelum ada peringkat
- AMPK Promove Expressão Gênica de GLUT 4 em Humanos. MCGGE (2008)Dokumen8 halamanAMPK Promove Expressão Gênica de GLUT 4 em Humanos. MCGGE (2008)Anderson Ranieri MassahudBelum ada peringkat
- Mutations in The EGFR Kinase Domain Mediate STAT3 Activation Via IL-6 Production in Human Lung AdenocarcinomasDokumen11 halamanMutations in The EGFR Kinase Domain Mediate STAT3 Activation Via IL-6 Production in Human Lung AdenocarcinomasNilabh RanjanBelum ada peringkat
- Farnesyltransferase InhibitorDokumen4 halamanFarnesyltransferase InhibitorLê Khánh ToànBelum ada peringkat
- Effects of Propolis and Phenolic Acids On Triple-NDokumen14 halamanEffects of Propolis and Phenolic Acids On Triple-Nriyadh jizan ُegyptian-boys-sectionBelum ada peringkat
- 2013-Cancer Research-Resistance To Irreversible EGF Receptor Tyrosine Kinase Inhibitors Through A Multistep Mechanism Involving The IGF1R PathwayDokumen10 halaman2013-Cancer Research-Resistance To Irreversible EGF Receptor Tyrosine Kinase Inhibitors Through A Multistep Mechanism Involving The IGF1R Pathwayzhe zhBelum ada peringkat
- EPZ015666, A SAM-cooperative Selective Inhibitor of PRMT5 With in Vivo and in Vitro Potency in MCL ModelsDokumen10 halamanEPZ015666, A SAM-cooperative Selective Inhibitor of PRMT5 With in Vivo and in Vitro Potency in MCL ModelsMatthew FengBelum ada peringkat
- BPA + TMF-a Induce Th2 Promoting DCsDokumen8 halamanBPA + TMF-a Induce Th2 Promoting DCsshweta pasiBelum ada peringkat
- The Role of Immune Checkpoints in Immunity and CancerDokumen8 halamanThe Role of Immune Checkpoints in Immunity and Cancernikos tsatsakisBelum ada peringkat
- tmpD101 TMPDokumen10 halamantmpD101 TMPFrontiersBelum ada peringkat
- Jcinvest00043 0362Dokumen8 halamanJcinvest00043 0362api-26848049Belum ada peringkat
- tmp90FE TMPDokumen10 halamantmp90FE TMPFrontiersBelum ada peringkat
- Irinotecan: Mechanisms of Tumor Resistance and Novel Strategies For Modulating Its ActivityDokumen11 halamanIrinotecan: Mechanisms of Tumor Resistance and Novel Strategies For Modulating Its ActivityCindyGaniBelum ada peringkat
- Bcr-Abl Tyrosine-Kinase Inhibitor: HistoryDokumen31 halamanBcr-Abl Tyrosine-Kinase Inhibitor: HistoryHemansu PathakBelum ada peringkat
- Plakat EMBO 2011Dokumen1 halamanPlakat EMBO 2011Magdalena PruszkoBelum ada peringkat
- Autophagy PapersDokumen7 halamanAutophagy PapersKudumarBelum ada peringkat
- p53 (Tumor Suppressor Gene) : Presented By: Saeed Rashid Presented To: Sir YasirDokumen25 halamanp53 (Tumor Suppressor Gene) : Presented By: Saeed Rashid Presented To: Sir YasirSaeed RashidBelum ada peringkat
- Structure-Activity Relationships of Synthetic Analogs of (-) - Epigallocatechin-3-Gallate As Proteasome InhibitorsDokumen12 halamanStructure-Activity Relationships of Synthetic Analogs of (-) - Epigallocatechin-3-Gallate As Proteasome InhibitorsGhibli GinBelum ada peringkat
- Role of of Protein Tyrosine Phosphatase Receptor Type in Chronic Myeloid LeukemiaDokumen3 halamanRole of of Protein Tyrosine Phosphatase Receptor Type in Chronic Myeloid Leukemiapatologi klinikBelum ada peringkat
- Sushi Domain LiewDokumen7 halamanSushi Domain Liewetincelle5690Belum ada peringkat
- Application of Recombinant DNATechnologies On Sub-Cloning of Transcriptional Co-FactorDokumen6 halamanApplication of Recombinant DNATechnologies On Sub-Cloning of Transcriptional Co-FactorInternational Journal of Innovative Science and Research TechnologyBelum ada peringkat
- European Journal of Medicinal Chemistry: Lingyi Sun, Hriday Bera, Wai Keung ChuiDokumen11 halamanEuropean Journal of Medicinal Chemistry: Lingyi Sun, Hriday Bera, Wai Keung ChuiViviana TorresBelum ada peringkat
- A Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)Dokumen38 halamanA Patent Review of Arginine Methyltransferase Inhibitors (2010-2018)shamimBelum ada peringkat
- HDACIs BET Protein Part IIIDokumen22 halamanHDACIs BET Protein Part IIIThomas hearnsBelum ada peringkat
- Artigo OncoDokumen14 halamanArtigo OncoCatarina LuísBelum ada peringkat
- Tanida 2012Dokumen9 halamanTanida 2012David Felipe Rojas DiazBelum ada peringkat
- Molecular BiologyDokumen44 halamanMolecular BiologyYasmin BalochBelum ada peringkat
- MCB00436-22-Merged - PDF MergedDokumen47 halamanMCB00436-22-Merged - PDF MergedHector Ivan Saldivar CeronBelum ada peringkat
- Novel Sensitizing Agents for Therapeutic Anti-EGFR AntibodiesDari EverandNovel Sensitizing Agents for Therapeutic Anti-EGFR AntibodiesShi HuBelum ada peringkat
- InmunosupresoresDokumen8 halamanInmunosupresoresISABEL SOFIA CABARCAS COGOLLOBelum ada peringkat
- Gamma-Tocopherol or Combinations of Vitamin E Forms Induce Cell Death in Human Prostate Cancer Cells. Jiang, Et Al., 2004.Dokumen6 halamanGamma-Tocopherol or Combinations of Vitamin E Forms Induce Cell Death in Human Prostate Cancer Cells. Jiang, Et Al., 2004.ytreffalBelum ada peringkat
- Artículo+Dokumen10 halamanArtículo+king130Belum ada peringkat
- Crozza 2020-BioRxiv-ENPP1 Inhibitors - ReviewDokumen37 halamanCrozza 2020-BioRxiv-ENPP1 Inhibitors - ReviewBrahmam PujalaBelum ada peringkat
- 10 1016@j Ejmech 2019 111926Dokumen56 halaman10 1016@j Ejmech 2019 111926InbalBelum ada peringkat
- Research Article: Metformin Induces Cell Cycle Arrest and Apoptosis in Drug-Resistant Leukemia CellsDokumen12 halamanResearch Article: Metformin Induces Cell Cycle Arrest and Apoptosis in Drug-Resistant Leukemia Cellsnurfitriani runduBelum ada peringkat
- Dihydroflavonol BB-1, An Extract of Natural Plant Blumea Balsamifera, Abrogates TRAIL Resistance in Leukemia CellsDokumen11 halamanDihydroflavonol BB-1, An Extract of Natural Plant Blumea Balsamifera, Abrogates TRAIL Resistance in Leukemia Cellshermila nopiantiBelum ada peringkat
- Jurnal BioselDokumen15 halamanJurnal BioselRegita cahyani priscalisyiBelum ada peringkat
- EDokumen5 halamanEJohn HolmanBelum ada peringkat
- Marketing MixDokumen1 halamanMarketing MixJohn HolmanBelum ada peringkat
- HDokumen3 halamanHJohn HolmanBelum ada peringkat
- GullibleDokumen4 halamanGullibleJohn HolmanBelum ada peringkat
- Dogon SleepDokumen1 halamanDogon SleepJohn HolmanBelum ada peringkat
- OutkastDokumen12 halamanOutkastJohn HolmanBelum ada peringkat
- Make MoneyDokumen6 halamanMake MoneyJohn HolmanBelum ada peringkat
- DDokumen3 halamanDJohn HolmanBelum ada peringkat
- DDokumen3 halamanDJohn HolmanBelum ada peringkat
- Make Money 2Dokumen5 halamanMake Money 2John HolmanBelum ada peringkat
- ADokumen4 halamanAJohn HolmanBelum ada peringkat
- Youtube 5Dokumen4 halamanYoutube 5John HolmanBelum ada peringkat
- CDokumen5 halamanCJohn HolmanBelum ada peringkat
- Make Money 3Dokumen4 halamanMake Money 3John HolmanBelum ada peringkat
- Youtube 9Dokumen16 halamanYoutube 9John HolmanBelum ada peringkat
- Science 3Dokumen4 halamanScience 3John HolmanBelum ada peringkat
- Science 4Dokumen3 halamanScience 4John HolmanBelum ada peringkat
- Spirit 4Dokumen5 halamanSpirit 4John HolmanBelum ada peringkat
- Siegler Etal InpressPsySciDokumen30 halamanSiegler Etal InpressPsySciJohn HolmanBelum ada peringkat
- Science 1Dokumen3 halamanScience 1John HolmanBelum ada peringkat
- Youtube 8Dokumen6 halamanYoutube 8John HolmanBelum ada peringkat
- Youtube 7Dokumen3 halamanYoutube 7John HolmanBelum ada peringkat
- Youtube 7Dokumen3 halamanYoutube 7John HolmanBelum ada peringkat
- Science 2Dokumen3 halamanScience 2John HolmanBelum ada peringkat
- yOUTUBE 2Dokumen4 halamanyOUTUBE 2John HolmanBelum ada peringkat
- Youtube 3Dokumen3 halamanYoutube 3John HolmanBelum ada peringkat
- Youtube 5Dokumen4 halamanYoutube 5John HolmanBelum ada peringkat
- yOUTUBE 2Dokumen4 halamanyOUTUBE 2John HolmanBelum ada peringkat
- Youtube 4Dokumen5 halamanYoutube 4John HolmanBelum ada peringkat
- Kinase InhibitorDokumen85 halamanKinase InhibitorSowjanya NekuriBelum ada peringkat
- Bavetsias Et Al-2015-Frontiers in OncologyDokumen10 halamanBavetsias Et Al-2015-Frontiers in OncologyJubairBelum ada peringkat
- Acute Lymphoblastic Leukemia in The Older Adult: Ibrahim Aldoss, MD Stephen J. Forman, MD and Vinod Pullarkat, MDDokumen10 halamanAcute Lymphoblastic Leukemia in The Older Adult: Ibrahim Aldoss, MD Stephen J. Forman, MD and Vinod Pullarkat, MDHugo HectorBelum ada peringkat
- Drug Resistance and Drug Synergism - Chapter 7Dokumen40 halamanDrug Resistance and Drug Synergism - Chapter 7Shaun李好Belum ada peringkat
- Chronic Myeloid LeukaemiaDokumen14 halamanChronic Myeloid LeukaemiaAnggie Pradetya MaharaniBelum ada peringkat
- Mechanisms of Resistance To Imatinib and Second-GenerationDokumen10 halamanMechanisms of Resistance To Imatinib and Second-GenerationRiyadh Z. MawloodBelum ada peringkat
- Biology Investigatory Project CMLDokumen16 halamanBiology Investigatory Project CMLABHISHEK AjithkumarBelum ada peringkat
- Investor Presentation - Update On NCE and NDDS Programs (Company Update)Dokumen61 halamanInvestor Presentation - Update On NCE and NDDS Programs (Company Update)Shyam SunderBelum ada peringkat