Anda mungkin juga menyukai
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Dist - Propor.danfoss PVG32Dokumen136 halamanDist - Propor.danfoss PVG32Michal BujaraBelum ada peringkat
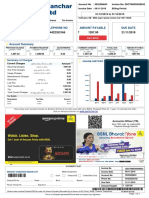
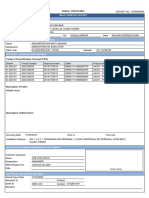
- BSNL BillDokumen3 halamanBSNL BillKaushik GurunathanBelum ada peringkat
- BSRM Ultima BrochureDokumen2 halamanBSRM Ultima BrochuresaifuzzamanBelum ada peringkat
- Libya AIP Part1Dokumen145 halamanLibya AIP Part1Hitham Ghwiel100% (1)
- Optical Transport Network SwitchingDokumen16 halamanOptical Transport Network SwitchingNdambuki DicksonBelum ada peringkat
- Sharat Babu Digumarti Vs State, Govt. of NCT of Delhi (Bazee - Com Case, Appeal) - Information Technology ActDokumen17 halamanSharat Babu Digumarti Vs State, Govt. of NCT of Delhi (Bazee - Com Case, Appeal) - Information Technology ActRavish Rana100% (1)
- Journal of Molecular LiquidsDokumen11 halamanJournal of Molecular LiquidsDennys MacasBelum ada peringkat
- Revenue Memorandum Circular No. 55-2016: For ExampleDokumen2 halamanRevenue Memorandum Circular No. 55-2016: For ExampleFedsBelum ada peringkat
- ManualDokumen50 halamanManualspacejung50% (2)
- Fuentes v. Office of The Ombudsman - MindanaoDokumen6 halamanFuentes v. Office of The Ombudsman - MindanaoJ. JimenezBelum ada peringkat
- Documentos de ExportaçãoDokumen17 halamanDocumentos de ExportaçãoZineBelum ada peringkat
- Flight Data Recorder Rule ChangeDokumen7 halamanFlight Data Recorder Rule ChangeIgnacio ZupaBelum ada peringkat
- SCIENCE 11 WEEK 6c - Endogenic ProcessDokumen57 halamanSCIENCE 11 WEEK 6c - Endogenic ProcessChristine CayosaBelum ada peringkat
- Tenses English Grammar PresentationDokumen14 halamanTenses English Grammar PresentationMaz Gedi60% (5)
- Art Integrated ProjectDokumen14 halamanArt Integrated ProjectSreeti GangulyBelum ada peringkat
- What Is The PCB Shelf Life Extending The Life of PCBsDokumen9 halamanWhat Is The PCB Shelf Life Extending The Life of PCBsjackBelum ada peringkat
- Measuring Road Roughness by Static Level Method: Standard Test Method ForDokumen6 halamanMeasuring Road Roughness by Static Level Method: Standard Test Method ForDannyChaconBelum ada peringkat
- APJ Abdul Kalam Success StoryDokumen1 halamanAPJ Abdul Kalam Success StorySanjaiBelum ada peringkat
- Final - Anarchy One-Sheet Sell SheetDokumen2 halamanFinal - Anarchy One-Sheet Sell SheetMaddanie WijayaBelum ada peringkat
- Experiment 2 HORSEPOWER EFFICIENCY GEAR RATIO AND SPEED RATIODokumen10 halamanExperiment 2 HORSEPOWER EFFICIENCY GEAR RATIO AND SPEED RATIOJake Polo SantiagoBelum ada peringkat
- Nat Steel BREGENEPD000379Dokumen16 halamanNat Steel BREGENEPD000379Batu GajahBelum ada peringkat
- Communicating Value - PatamilkaDokumen12 halamanCommunicating Value - PatamilkaNeha ArumallaBelum ada peringkat
- The Anti-PaladinDokumen9 halamanThe Anti-PaladinBobbyBelum ada peringkat
- Army Aviation Digest - Apr 1971Dokumen68 halamanArmy Aviation Digest - Apr 1971Aviation/Space History LibraryBelum ada peringkat
- Extraction of Mangiferin From Mangifera Indica L. LeavesDokumen7 halamanExtraction of Mangiferin From Mangifera Indica L. LeavesDaniel BartoloBelum ada peringkat
- LP Week 8Dokumen4 halamanLP Week 8WIBER ChapterLampungBelum ada peringkat
- DBM Uv W ChartDokumen2 halamanDBM Uv W ChartEddie FastBelum ada peringkat
- Blackbook 2Dokumen94 halamanBlackbook 2yogesh kumbharBelum ada peringkat
- Investigation Data FormDokumen1 halamanInvestigation Data Formnildin danaBelum ada peringkat
- Docket - CDB Batu GajahDokumen1 halamanDocket - CDB Batu Gajahfatin rabiatul adawiyahBelum ada peringkat