Anda mungkin juga menyukai
- Codigodebarras 100527231907 Phpapp02Dokumen53 halamanCodigodebarras 100527231907 Phpapp02sena virtualBelum ada peringkat
- RobóticaDokumen14 halamanRobóticaClaudia DiazBelum ada peringkat
- Codigodebarras 140905090134 Phpapp01Dokumen19 halamanCodigodebarras 140905090134 Phpapp01Claudia DiazBelum ada peringkat
- Cdigosbarra 140225133431 Phpapp02Dokumen6 halamanCdigosbarra 140225133431 Phpapp02Claudia DiazBelum ada peringkat
- Cirugarobtica 140320113050 Phpapp01Dokumen20 halamanCirugarobtica 140320113050 Phpapp01Claudia DiazBelum ada peringkat
- Atlas HematologiaDokumen83 halamanAtlas HematologiaGamaliel Canté HdezBelum ada peringkat
- Diapositivasdeltrabajodelarobotica 121113113104 Phpapp02Dokumen10 halamanDiapositivasdeltrabajodelarobotica 121113113104 Phpapp02claudia palominoBelum ada peringkat
- Desprendimientodevtreo 151016080054 Lva1 App6892Dokumen31 halamanDesprendimientodevtreo 151016080054 Lva1 App6892Claudia DiazBelum ada peringkat
- Cdigodebarras 101101090022 Phpapp02Dokumen2 halamanCdigodebarras 101101090022 Phpapp02Claudia DiazBelum ada peringkat
- Davinci 131025023058 Phpapp02Dokumen17 halamanDavinci 131025023058 Phpapp02claudia palominoBelum ada peringkat
- Conceptosbasicosderobotica 120428120714 Phpapp01Dokumen28 halamanConceptosbasicosderobotica 120428120714 Phpapp01Jonathan BJBelum ada peringkat
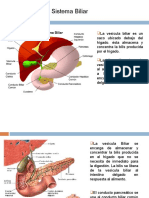
- Anatomía Del Sistema BiliarDokumen25 halamanAnatomía Del Sistema Biliarclaudia palominoBelum ada peringkat
- Etapas de La CoagulacionDokumen7 halamanEtapas de La CoagulacionClaudia DiazBelum ada peringkat
- Sociograma AvanceDokumen1 halamanSociograma AvanceClaudia DiazBelum ada peringkat
- Esunbeneficioelrobot Da Vincienoperacionesdela PrstataDokumen5 halamanEsunbeneficioelrobot Da Vincienoperacionesdela PrstataClaudia DiazBelum ada peringkat
- Trombocitemia esencial: definición, fisiopatología, manifestaciones clínicas y tratamientoDokumen17 halamanTrombocitemia esencial: definición, fisiopatología, manifestaciones clínicas y tratamientoClaudia DiazBelum ada peringkat
- Esunbeneficioelrobot Da Vincienoperacionesdela PrstataDokumen5 halamanEsunbeneficioelrobot Da Vincienoperacionesdela PrstataClaudia DiazBelum ada peringkat
- Expo AmbientalDokumen47 halamanExpo AmbientalClaudia DiazBelum ada peringkat
- Punto en XDokumen4 halamanPunto en XNoel Saúl Argüello SánchezBelum ada peringkat
- Sutura Quirurgica Simple y ContinuaDokumen104 halamanSutura Quirurgica Simple y ContinuaClaudia DiazBelum ada peringkat
- Diapositivas de La Historia de La AnatomiaDokumen120 halamanDiapositivas de La Historia de La AnatomiaClaudia DiazBelum ada peringkat
- Calvo AnemiaDokumen22 halamanCalvo AnemiaKelly Jessica Fernandez LandeoBelum ada peringkat
- Fisiopatología del tromboembolismo pulmonarDokumen6 halamanFisiopatología del tromboembolismo pulmonarClaudia DiazBelum ada peringkat
- Expo ListaDokumen11 halamanExpo ListaClaudia DiazBelum ada peringkat
- Pato Genetica PDFDokumen14 halamanPato Genetica PDFkarla7barrenecheaBelum ada peringkat
- Anemia Por Enfermedades CronicasDokumen17 halamanAnemia Por Enfermedades CronicasClaudia DiazBelum ada peringkat
- Tromboembolismopulmonar 150813210050 Lva1 App6892 PDFDokumen70 halamanTromboembolismopulmonar 150813210050 Lva1 App6892 PDFClaudia DiazBelum ada peringkat
- Trombosis VenosaprofundaDokumen4 halamanTrombosis VenosaprofundaClaudia DiazBelum ada peringkat
- Trombosisvenosaprofunda 120824225908 Phpapp02Dokumen33 halamanTrombosisvenosaprofunda 120824225908 Phpapp02Claudia DiazBelum ada peringkat
- Tromboembolismopulmonar 150813210050 Lva1 App6892Dokumen70 halamanTromboembolismopulmonar 150813210050 Lva1 App6892Claudia DiazBelum ada peringkat
- TMVDokumen18 halamanTMVNestor JuarezBelum ada peringkat
- Flora MicrobianaDokumen81 halamanFlora MicrobianaNilson NavarroBelum ada peringkat
- Unidad IV Tema 3 Bioinformatica Clinica REVISADODokumen33 halamanUnidad IV Tema 3 Bioinformatica Clinica REVISADOGraciela Estrada PortocarreroBelum ada peringkat
- Tema 19. Ingeniería Genética - MioDokumen26 halamanTema 19. Ingeniería Genética - MioLeticia Foncubierta100% (1)
- Práctica 3 PUREZA ADNDokumen3 halamanPráctica 3 PUREZA ADNHernández Murillo Alaír José JuanBelum ada peringkat
- BioquímicaDokumen9 halamanBioquímicaFernanda NuñezBelum ada peringkat
- Ácidos Nucleicos (Modo de Compatibilidad)Dokumen114 halamanÁcidos Nucleicos (Modo de Compatibilidad)Alejo Diez PatiñoBelum ada peringkat
- Dogma Central de La Biología..Dokumen5 halamanDogma Central de La Biología..Edgar Humberto Crecencio HernándezBelum ada peringkat
- Presentacion Adn y ArnDokumen12 halamanPresentacion Adn y Arnlinda yurani izquierdo valoyBelum ada peringkat
- Extracción ADN diagnóstico molecularDokumen4 halamanExtracción ADN diagnóstico molecularMARGARETH JULIETH PEÑA JARABABelum ada peringkat
- Navarra Biologia-2019Dokumen8 halamanNavarra Biologia-2019Paula Gonzalez MesaBelum ada peringkat
- Pract. #2 Extracción de ADNDokumen5 halamanPract. #2 Extracción de ADNSandy BernalBelum ada peringkat
- Prueba Diagnóstica Biología 1Dokumen3 halamanPrueba Diagnóstica Biología 1Andrea Catuto100% (2)
- ELECTROFORESISDokumen26 halamanELECTROFORESISchonita_54100% (1)
- Taller No. 2 Microbiología 2 2020Dokumen8 halamanTaller No. 2 Microbiología 2 2020Liliana Cely UscateguiBelum ada peringkat
- Apuntes de MicrobiologíaDokumen74 halamanApuntes de Microbiologíaamanda grimaldoBelum ada peringkat
- El Reloj Biológico Del EnvejecimientoDokumen8 halamanEl Reloj Biológico Del EnvejecimientoJavier Perez DrizBelum ada peringkat
- Cromosomas PolitenicosDokumen3 halamanCromosomas PolitenicosMariana JiménezBelum ada peringkat
- Informe Proceso InflamatorioDokumen11 halamanInforme Proceso InflamatorioMoises RodriguezBelum ada peringkat
- Formas de Reproducción BacterianaDokumen5 halamanFormas de Reproducción BacterianaHABNER123456Belum ada peringkat
- Fisiopatologia, Unidad 1Dokumen38 halamanFisiopatologia, Unidad 1Diego PintoBelum ada peringkat
- ¿Que Nos Enseña Sobre La Cultura Humana Los Primates Vivos No Humanos?Dokumen9 halaman¿Que Nos Enseña Sobre La Cultura Humana Los Primates Vivos No Humanos?Angela ParraBelum ada peringkat
- Marcadores moleculares DNADokumen60 halamanMarcadores moleculares DNALuis Ricardo Rodriguez VaresBelum ada peringkat
- Los 12 Filamentos Del ADNDokumen3 halamanLos 12 Filamentos Del ADNFederico GaleanoBelum ada peringkat
- Antineoplasicos PDFDokumen38 halamanAntineoplasicos PDFORACIO100% (1)
- Ciencias Naturales Primer Ano de BachilleratoDokumen31 halamanCiencias Naturales Primer Ano de BachilleratoAlexandra TurciosBelum ada peringkat
- Traduccion Capitulo 19Dokumen28 halamanTraduccion Capitulo 19Jennifer LizarazoBelum ada peringkat
- Importancia de La Prueba Del ADN en La Impartición de Justicia Trabajo Invest 1Dokumen35 halamanImportancia de La Prueba Del ADN en La Impartición de Justicia Trabajo Invest 1Celestino Valverde SoteloBelum ada peringkat
- Ahumada, P. (2015) - ¿Por Qué Sobrevive El Sistema CapitalistaDokumen166 halamanAhumada, P. (2015) - ¿Por Qué Sobrevive El Sistema CapitalistaSebastian33% (3)
- En La Frontera de La VidaDokumen35 halamanEn La Frontera de La Vidaperfil100realnofake0% (1)