Anda mungkin juga menyukai
- Insulin Resistance: Childhood Precursors of Adult DiseaseDari EverandInsulin Resistance: Childhood Precursors of Adult DiseasePhilip S. ZeitlerBelum ada peringkat
- Sex Steroids and Insulin Resistance: Callum LIVINGSTONE and Mary COLLISONDokumen16 halamanSex Steroids and Insulin Resistance: Callum LIVINGSTONE and Mary COLLISONputrefactio6669Belum ada peringkat
- Prediction of Type 2 Diabetes: A Natural History PerspectiveDokumen9 halamanPrediction of Type 2 Diabetes: A Natural History Perspectiveandree_zz4890Belum ada peringkat
- Literature Review On Type 2 Diabetes MellitusDokumen7 halamanLiterature Review On Type 2 Diabetes Mellituscmppcmwgf100% (1)
- Cons EsusDokumen10 halamanCons EsusΠέγκυΓρηγοροπούλουBelum ada peringkat
- Diabetes Mellitus Type 2 Research PaperDokumen6 halamanDiabetes Mellitus Type 2 Research Paperntjjkmrhf100% (1)
- Endocrine Week 2Dokumen18 halamanEndocrine Week 2Clauvinna Lie FiscaBelum ada peringkat
- Pathophysiology of Insulin ResistanceDokumen8 halamanPathophysiology of Insulin ResistanceCamilo VidalBelum ada peringkat
- Pa Tho Genesis of Diabetes MellitusDokumen6 halamanPa Tho Genesis of Diabetes MellitusBram WongsoBelum ada peringkat
- Obesity Research - 2012 - VozarovaDokumen5 halamanObesity Research - 2012 - VozarovaKarla SanchezBelum ada peringkat
- Insulin Resistance PHD ThesisDokumen7 halamanInsulin Resistance PHD Thesisjessicaoatisneworleans100% (2)
- S105 FullDokumen7 halamanS105 FullNurul HidayatiBelum ada peringkat
- KNH 411 - Case Study 22Dokumen17 halamanKNH 411 - Case Study 22api-301118772100% (3)
- Literature Review On Insulin ResistanceDokumen8 halamanLiterature Review On Insulin Resistanceyjdzpccnd100% (1)
- Humans With Type 2 Diabetes: - Cell Deficit and Increased - Cell Apoptosis inDokumen9 halamanHumans With Type 2 Diabetes: - Cell Deficit and Increased - Cell Apoptosis ingrilo604Belum ada peringkat
- Diabetes AssignmentDokumen8 halamanDiabetes AssignmentMildred Obeng AgyeiwaaBelum ada peringkat
- Medication-Induced Diabetes MellitusDokumen6 halamanMedication-Induced Diabetes MellitusAmjadBelum ada peringkat
- (1479683X - European Journal of Endocrinology) Subcutaneous Adipocytes From Obese Hyperinsulinemic Women With Polycystic Ovary Syndrome Exhibit Normal Insulin Sensitivity But ReducedDokumen5 halaman(1479683X - European Journal of Endocrinology) Subcutaneous Adipocytes From Obese Hyperinsulinemic Women With Polycystic Ovary Syndrome Exhibit Normal Insulin Sensitivity But ReduceddfdfffffBelum ada peringkat
- PCOS and Metabolic SyndromeDokumen21 halamanPCOS and Metabolic SyndromeHAVIZ YUADBelum ada peringkat
- Diabetes Case StudyDokumen9 halamanDiabetes Case Studyapi-346620455Belum ada peringkat
- Nutrient Regulation of InsulinDokumen16 halamanNutrient Regulation of InsulinNickolasMarllesBelum ada peringkat
- Case 16 QuestionsDokumen10 halamanCase 16 Questionsapi-532124328Belum ada peringkat
- 2 Paginas Art 5 CommentaryDokumen2 halaman2 Paginas Art 5 CommentaryNilton SantibanezBelum ada peringkat
- Gluco TypesDokumen23 halamanGluco TypesSuresh giriBelum ada peringkat
- Diabetes in Elderly Adults: Graydon S. Meneilly and Daniel TessierDokumen9 halamanDiabetes in Elderly Adults: Graydon S. Meneilly and Daniel Tessierdita prameswariBelum ada peringkat
- Garber 2007Dokumen6 halamanGarber 2007Diana SamuezaBelum ada peringkat
- Foreword: Prof. Jacek MalejczykDokumen1 halamanForeword: Prof. Jacek MalejczykFazalJaturieBelum ada peringkat
- Review ArticleDokumen9 halamanReview ArticleottohasibuBelum ada peringkat
- VHRM - 6098 - Inhaled Insulin - An Overview of A Novel Form of Inhaled Ins - 011310Dokumen12 halamanVHRM - 6098 - Inhaled Insulin - An Overview of A Novel Form of Inhaled Ins - 011310Rubana Reaz TanaBelum ada peringkat
- Sumber Resist Insulin (English)Dokumen15 halamanSumber Resist Insulin (English)Muhammad Amri KautsarBelum ada peringkat
- Diabetes Mellitus in Children (Pedi Clinics of N. America Vol 52, No 6) WW PDFDokumen275 halamanDiabetes Mellitus in Children (Pedi Clinics of N. America Vol 52, No 6) WW PDFEliMihaelaBelum ada peringkat
- Research Paper On Diabetes Mellitus Type 2Dokumen4 halamanResearch Paper On Diabetes Mellitus Type 2gvzrg8jy100% (1)
- The Cellular Fate of Glucose and Its Relevance In-1-16Dokumen16 halamanThe Cellular Fate of Glucose and Its Relevance In-1-16Yariela Amador MendietaBelum ada peringkat
- Cellular Mechanisms of Insulin Resistance.Dokumen6 halamanCellular Mechanisms of Insulin Resistance.João AndradeBelum ada peringkat
- NASH in ChildrenDokumen10 halamanNASH in ChildrendrtpkBelum ada peringkat
- MoonikaDokumen5 halamanMoonikaDinda SaviraBelum ada peringkat
- Gene Therapy Uncovers Role of Leptin in DiabetesDokumen7 halamanGene Therapy Uncovers Role of Leptin in Diabetesjinsung parkBelum ada peringkat
- Title of The Research: Diabetes Mellitus By: Abdelrahman Gamal Course Title: Pharmacology Academic Year: Fourth 2019-2020Dokumen11 halamanTitle of The Research: Diabetes Mellitus By: Abdelrahman Gamal Course Title: Pharmacology Academic Year: Fourth 2019-2020LAZKILLERBelum ada peringkat
- Case Study Analysis of Type I Diabetes MellitusDokumen9 halamanCase Study Analysis of Type I Diabetes MellitusJyrelle Rhyo YparraguirreBelum ada peringkat
- Diabetes Research Paper IntroductionDokumen9 halamanDiabetes Research Paper Introductiongz7vxzyz100% (1)
- Obesity, Insulin Resistance, and Cardiovascular DiseaseDokumen17 halamanObesity, Insulin Resistance, and Cardiovascular DiseaseKaro GarrydoBelum ada peringkat
- ResponseConsensusReportAntipsychoticsMetabolicIssuesLetter CITROME DiabCare2004Dokumen5 halamanResponseConsensusReportAntipsychoticsMetabolicIssuesLetter CITROME DiabCare2004Leslie CitromeBelum ada peringkat
- Non Alcoholic Fatty Liver Disease and Metabolic Syndrome: Invited ArticleDokumen11 halamanNon Alcoholic Fatty Liver Disease and Metabolic Syndrome: Invited ArticleBayu GemilangBelum ada peringkat
- Paper 8688Dokumen14 halamanPaper 8688IJARSCT JournalBelum ada peringkat
- Regenerative medicine approaches for diabetes treatmentDokumen10 halamanRegenerative medicine approaches for diabetes treatmentAlex PlicBelum ada peringkat
- Article - Ghrelin and Leptin in Insulin Resistance (CHECKED)Dokumen6 halamanArticle - Ghrelin and Leptin in Insulin Resistance (CHECKED)pixoguiasBelum ada peringkat
- Metabolic Syndrome and Diabetes: A Bidirectional RelationshipDokumen6 halamanMetabolic Syndrome and Diabetes: A Bidirectional RelationshipKarla MonsalvoBelum ada peringkat
- Effects of Chronic Protein-Calorie Malnutrition On The KidneyDokumen13 halamanEffects of Chronic Protein-Calorie Malnutrition On The KidneyAgnihotram GopinathBelum ada peringkat
- Gestational Diabetes IrDokumen10 halamanGestational Diabetes IrChikezie OnwukweBelum ada peringkat
- NIH Public AccessDokumen23 halamanNIH Public AccesssiscupBelum ada peringkat
- Resistencia A La Insulina Madre y FetoDokumen10 halamanResistencia A La Insulina Madre y FetoNorma AlvarezBelum ada peringkat
- Insulin Resistance ThesisDokumen7 halamanInsulin Resistance Thesismichellemaderowichitafalls100% (2)
- Metabolic Evaluation of Young Women With Congenital Adrenal HyperplasiaDokumen7 halamanMetabolic Evaluation of Young Women With Congenital Adrenal HyperplasiaOnysapto PramanaBelum ada peringkat
- Type1 DiabetesDokumen7 halamanType1 DiabetesbettyborbleBelum ada peringkat
- DM Tipe 1Dokumen6 halamanDM Tipe 1Mutiara Kristiani PutriBelum ada peringkat
- Insulin injection and pump options for diabetes patientsDokumen7 halamanInsulin injection and pump options for diabetes patientsSourav DasBelum ada peringkat
- Gestational Diabetes Mellitus: Science in MedicineDokumen7 halamanGestational Diabetes Mellitus: Science in MedicineNatalia_p_mBelum ada peringkat
- Maturity Onset Diabetes of The Young: Review ArticleDokumen8 halamanMaturity Onset Diabetes of The Young: Review ArticleMartha VillacisBelum ada peringkat
- Costa Rica GlucoseDokumen91 halamanCosta Rica GlucoseAnthony HarderBelum ada peringkat
- Fatty Pancreas Clinical ImplicationsDokumen6 halamanFatty Pancreas Clinical ImplicationsEngin ALTINTASBelum ada peringkat
- TicketDokumen1 halamanTicketIke Mahlida PBelum ada peringkat
- 705 FullDokumen13 halaman705 FullIke Mahlida PBelum ada peringkat
- APLIKASI ANALISIS KETAHANAN PADA DATA ANAK PUTUS SEKOLAH (Application of Survival Analysis On The Data of Drop Out Students) Nurul Astuty YensyDokumen14 halamanAPLIKASI ANALISIS KETAHANAN PADA DATA ANAK PUTUS SEKOLAH (Application of Survival Analysis On The Data of Drop Out Students) Nurul Astuty YensyIke Mahlida PBelum ada peringkat
- Repair of Pancreatic P-Cells: A Relevant Phenomenon in Early IDDM?Dokumen9 halamanRepair of Pancreatic P-Cells: A Relevant Phenomenon in Early IDDM?Ike Mahlida PBelum ada peringkat
- Carl Hollander SociogramDokumen19 halamanCarl Hollander SociogramIke Mahlida P100% (1)
- Types of Sentences PDFDokumen26 halamanTypes of Sentences PDFnaomi1586Belum ada peringkat
- Answer Key Regression 2.78Dokumen5 halamanAnswer Key Regression 2.78Ike Mahlida PBelum ada peringkat
- 2nd Brochure PDFDokumen2 halaman2nd Brochure PDFhappy855Belum ada peringkat
- CBTDokumen21 halamanCBTsavvy_as_98Belum ada peringkat
- 45 PDFDokumen8 halaman45 PDFChika FebrianiBelum ada peringkat
- Rhubarb April 2017Dokumen17 halamanRhubarb April 2017ISADD LietuvaBelum ada peringkat
- Vidas Troponin High Sensitive Ref#415386Dokumen1 halamanVidas Troponin High Sensitive Ref#415386Mike GesmundoBelum ada peringkat
- CefepimeDokumen2 halamanCefepimeMae Ann Bueno CastillonBelum ada peringkat
- 11th Zoology EM Practical NotesDokumen11 halaman11th Zoology EM Practical NotesBhavana Gopinath100% (1)
- 3 Wives One Husband 2Dokumen508 halaman3 Wives One Husband 2relebohilelehasa2Belum ada peringkat
- DUR - Fracture of The Shaft Radius and UlnaDokumen55 halamanDUR - Fracture of The Shaft Radius and UlnaM. Abdurrahman Al-HaraaniBelum ada peringkat
- Lesson Plan in English 8 Q2 Module 1Dokumen8 halamanLesson Plan in English 8 Q2 Module 1Nicky SanchezBelum ada peringkat
- CPM14th DIABETES Food GuideDokumen21 halamanCPM14th DIABETES Food GuideXhiariwen Bayro0% (1)
- Seven Principles of Building ResilienceDokumen7 halamanSeven Principles of Building ResiliencemilosvblBelum ada peringkat
- Vivid I and Vivid Q Cardiovascular Ultrasound: SectorDokumen4 halamanVivid I and Vivid Q Cardiovascular Ultrasound: SectorShaikh Emran HossainBelum ada peringkat
- Montelukast SodiumDokumen10 halamanMontelukast SodiumikhasilBelum ada peringkat
- Aloe Vera-A Wonder Plant Its History, Cultivation and Medicinal Uses PDFDokumen4 halamanAloe Vera-A Wonder Plant Its History, Cultivation and Medicinal Uses PDFveronyk28Belum ada peringkat
- MRI Monitoring System Provides Safety and MobilityDokumen4 halamanMRI Monitoring System Provides Safety and MobilityAchiyat WinataBelum ada peringkat
- Statement On Controlled Organ Donation After Circulatory DeathDokumen10 halamanStatement On Controlled Organ Donation After Circulatory DeathHeidi ReyesBelum ada peringkat
- Closing The Gap 2012Dokumen127 halamanClosing The Gap 2012ABC News OnlineBelum ada peringkat
- Psycho Sexual 6ceu CourseDokumen78 halamanPsycho Sexual 6ceu CourseariizianaBelum ada peringkat
- 0nvkwxysn505jodxuxkr5z3v PDFDokumen2 halaman0nvkwxysn505jodxuxkr5z3v PDFAnjali Thomas50% (2)
- The Abc of Handover: E.D. Registrar A: Areas AllocationDokumen1 halamanThe Abc of Handover: E.D. Registrar A: Areas AllocationDiahNiatiBelum ada peringkat
- DHI Score PDFDokumen2 halamanDHI Score PDFAnonymous HNGH1oBelum ada peringkat
- Pharmacoeconomics - Part IIIDokumen29 halamanPharmacoeconomics - Part IIISima JabbariBelum ada peringkat
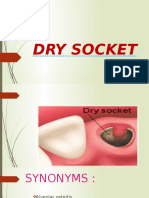
- Dry SocketDokumen20 halamanDry SocketHosuru SwethaBelum ada peringkat
- Standards OF Psychiatric Nursing PracticeDokumen15 halamanStandards OF Psychiatric Nursing Practicesameeram83Belum ada peringkat
- Chester V Afshar (2005) 1 A.C. 134Dokumen33 halamanChester V Afshar (2005) 1 A.C. 134V100% (1)
- Calculation SeatworkDokumen2 halamanCalculation SeatworkBenedict James BermasBelum ada peringkat
- Sinclair ch05 089-110Dokumen22 halamanSinclair ch05 089-110Shyamol BoseBelum ada peringkat
- Food and Exercise LogDokumen24 halamanFood and Exercise LogmhetfieldBelum ada peringkat
- Nexthealth - Semantic Web Sparks Evolution of Health 2.0 - A Road Map To Consumer-Centric HealthcareDokumen7 halamanNexthealth - Semantic Web Sparks Evolution of Health 2.0 - A Road Map To Consumer-Centric HealthcareMaarten den Braber100% (43)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDari EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionPenilaian: 4 dari 5 bintang4/5 (402)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisDari EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisPenilaian: 4 dari 5 bintang4/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDari EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedPenilaian: 5 dari 5 bintang5/5 (78)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDari EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityPenilaian: 4 dari 5 bintang4/5 (13)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDari EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeBelum ada peringkat
- The Comfort of Crows: A Backyard YearDari EverandThe Comfort of Crows: A Backyard YearPenilaian: 4.5 dari 5 bintang4.5/5 (23)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDari EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsPenilaian: 3.5 dari 5 bintang3.5/5 (3)
- Techniques Exercises And Tricks For Memory ImprovementDari EverandTechniques Exercises And Tricks For Memory ImprovementPenilaian: 4.5 dari 5 bintang4.5/5 (40)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDari EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsPenilaian: 5 dari 5 bintang5/5 (1)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsDari EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsPenilaian: 4.5 dari 5 bintang4.5/5 (169)
- The Obesity Code: Unlocking the Secrets of Weight LossDari EverandThe Obesity Code: Unlocking the Secrets of Weight LossPenilaian: 5 dari 5 bintang5/5 (4)
- The Ultimate Guide To Memory Improvement TechniquesDari EverandThe Ultimate Guide To Memory Improvement TechniquesPenilaian: 5 dari 5 bintang5/5 (34)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingDari EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingPenilaian: 5 dari 5 bintang5/5 (4)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDari EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDari EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsBelum ada peringkat
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisDari EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisPenilaian: 5 dari 5 bintang5/5 (8)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingDari EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingPenilaian: 3.5 dari 5 bintang3.5/5 (33)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Dari EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Penilaian: 4.5 dari 5 bintang4.5/5 (110)
- Summary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisDari EverandSummary: It Didn't Start with You: How Inherited Family Trauma Shapes Who We Are and How to End the Cycle By Mark Wolynn: Key Takeaways, Summary & AnalysisPenilaian: 5 dari 5 bintang5/5 (3)
- The Happiness Trap: How to Stop Struggling and Start LivingDari EverandThe Happiness Trap: How to Stop Struggling and Start LivingPenilaian: 4 dari 5 bintang4/5 (1)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeDari EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifePenilaian: 4.5 dari 5 bintang4.5/5 (253)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsDari EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsBelum ada peringkat
- The Tennis Partner: A Doctor's Story of Friendship and LossDari EverandThe Tennis Partner: A Doctor's Story of Friendship and LossPenilaian: 4.5 dari 5 bintang4.5/5 (4)