Anda mungkin juga menyukai
- Anatomie Topographique de La Région AxiliaireDokumen67 halamanAnatomie Topographique de La Région AxiliaireNadina TintiucBelum ada peringkat
- Cours Gestion CouleurDokumen9 halamanCours Gestion Couleurmikool19Belum ada peringkat
- Lien Mécanique PDFDokumen47 halamanLien Mécanique PDFPaweł Dziadoń (Manual Theory)Belum ada peringkat
- Dufour 1 Membre in FDokumen544 halamanDufour 1 Membre in Fcamilleblanc7Belum ada peringkat
- Fiche 12 - Gonio EpauleDokumen10 halamanFiche 12 - Gonio EpauleJedidi AmirBelum ada peringkat
- Biomécanique Du MS - Fiche de RévisionDokumen18 halamanBiomécanique Du MS - Fiche de RévisionCristina Mocan100% (1)
- Impertinente ostéopathie: Comprendre ses possibilités et ses limitesDari EverandImpertinente ostéopathie: Comprendre ses possibilités et ses limitesBelum ada peringkat
- Rudolf Steiner - Chaleur Et Matière - GA 321 PDFDokumen342 halamanRudolf Steiner - Chaleur Et Matière - GA 321 PDFbrunolorenzo111100% (2)
- Calcul Fondations ProfondesDokumen24 halamanCalcul Fondations ProfondesAhmed Ahmeed89% (9)
- 04-Le Potentiel de ReposDokumen34 halaman04-Le Potentiel de ReposBook Hunter100% (1)
- CH 1 - Gradients IoniquesDokumen12 halamanCH 1 - Gradients IoniquesTatiana Fontoura GermanoBelum ada peringkat
- 12 PeroxysomesDokumen5 halaman12 PeroxysomesImen Mestar100% (1)
- FrontalDokumen31 halamanFrontalBillel BenchihaBelum ada peringkat
- Electrophysiologie CardiaqueDokumen47 halamanElectrophysiologie CardiaqueLina Ray BenBelum ada peringkat
- 5 Cours Du Chapitre 1 L'électrophysiologieDokumen18 halaman5 Cours Du Chapitre 1 L'électrophysiologieAnnia VieBelum ada peringkat
- Paroi Abdominale PosterieureDokumen48 halamanParoi Abdominale PosterieureMrTriumphant100% (2)
- La Contraction MusculaireDokumen11 halamanLa Contraction MusculaireJihadi AbbouBelum ada peringkat
- ANATOMIE Appliquée RésuméDokumen20 halamanANATOMIE Appliquée RésuméSALMAN KHAYATIBelum ada peringkat
- Anatomie - VASCULARISATION DU MEMBRE INFERIEURDokumen21 halamanAnatomie - VASCULARISATION DU MEMBRE INFERIEURWladimir Jordan André100% (1)
- Articulations Du PoignetDokumen16 halamanArticulations Du PoignetJawad AymenBelum ada peringkat
- NeuroanatomieDokumen113 halamanNeuroanatomiePère ProsperBelum ada peringkat
- Biophysique - TD - Série 2-A PDFDokumen12 halamanBiophysique - TD - Série 2-A PDFAkram AzzouzBelum ada peringkat
- Tests Sacro-IliaquesDokumen41 halamanTests Sacro-IliaquesDr-Rodrigue HadchityBelum ada peringkat
- Les Epitheliums GlandulairesDokumen112 halamanLes Epitheliums GlandulairesAbdoulaziz SidilamineBelum ada peringkat
- Anatomie Biomecanique de CoudeDokumen8 halamanAnatomie Biomecanique de CoudeMohammed GartitBelum ada peringkat
- Muscles de La JambeDokumen13 halamanMuscles de La JambeBillel BenchihaBelum ada peringkat
- CORRECTION COLLE-JABOT-Homéostasie-01.10.22Dokumen6 halamanCORRECTION COLLE-JABOT-Homéostasie-01.10.22Yann legendreBelum ada peringkat
- Ostéopathie de La Cage ThoraciqueDokumen100 halamanOstéopathie de La Cage Thoraciquewhatsup100% (1)
- Electrophysiologie CardiaqueDokumen6 halamanElectrophysiologie CardiaqueMery SageBelum ada peringkat
- Histo Pcem2 Eb5534Dokumen86 halamanHisto Pcem2 Eb5534vitrina29Belum ada peringkat
- Ostéologie Du Membre SupérieurDokumen50 halamanOstéologie Du Membre SupérieurOli Via100% (2)
- Anatomie de L'oreille MoyenneDokumen19 halamanAnatomie de L'oreille MoyenneZineb BelmejdoubBelum ada peringkat
- Anatomie Palpatoire N°10Dokumen3 halamanAnatomie Palpatoire N°10ngpk9vzr5jBelum ada peringkat
- Tableau Des Nerfs CrâniensDokumen3 halamanTableau Des Nerfs CrâniensbihbibabiaBelum ada peringkat
- La BiomécaniqueDokumen13 halamanLa BiomécaniqueMeriem EL Hàmidii100% (1)
- QCM Nerv 2 PDFDokumen5 halamanQCM Nerv 2 PDFLara OliviaBelum ada peringkat
- Biomecanique Fonctionelle - 3 - ÉpauleDokumen43 halamanBiomecanique Fonctionelle - 3 - Épauleioana dark100% (1)
- Anat' Fonctionnelle CrânienneDokumen349 halamanAnat' Fonctionnelle Crâniennebooetienne30Belum ada peringkat
- Anatomie 1 Osteologie Arthrologie Et MylogieDokumen12 halamanAnatomie 1 Osteologie Arthrologie Et MylogieFlorine BaillietBelum ada peringkat
- Lhlaydi - AbdomenDokumen108 halamanLhlaydi - AbdomenYouness Khalfaoui100% (2)
- BiomecDokumen19 halamanBiomecKamel Fedaoui0% (1)
- EndocrinologieDokumen46 halamanEndocrinologieAhmedou EllaBelum ada peringkat
- QCM RadioprotectionDokumen14 halamanQCM Radioprotectionboussad1Belum ada peringkat
- Les Indispensables de La PACESDokumen4 halamanLes Indispensables de La PACESLucian NistorBelum ada peringkat
- Pratique Membre SupDokumen4 halamanPratique Membre SupBaptiste CarpentierBelum ada peringkat
- Generalites AnatDokumen12 halamanGeneralites AnatMaroua BensariBelum ada peringkat
- Neurophysiologie PDFDokumen140 halamanNeurophysiologie PDFPascal Kasero0% (1)
- Mémoire DIU 2015 Cheminon Neurodynamique Et CCDokumen74 halamanMémoire DIU 2015 Cheminon Neurodynamique Et CCmitroisergiuBelum ada peringkat
- Anatomie Biomecanique de L'epaule MODIFIERDokumen5 halamanAnatomie Biomecanique de L'epaule MODIFIERMohammed Gartit100% (1)
- Réticulum Endoplasmique OranDokumen7 halamanRéticulum Endoplasmique OranHaitam El OuahabiBelum ada peringkat
- ECG ExerciceDokumen30 halamanECG ExerciceAmineSlamaBelum ada peringkat
- Biophysique SommaireDokumen7 halamanBiophysique SommaireMoussaBelum ada peringkat
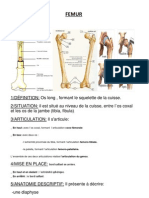
- FemurDokumen4 halamanFemurBesma SbBelum ada peringkat
- Testing MusculaireDokumen5 halamanTesting MusculaireNizar AkidBelum ada peringkat
- Poly - Pcem1-Theme 3 - by Med - TMSSDokumen142 halamanPoly - Pcem1-Theme 3 - by Med - TMSSjehane oussamaBelum ada peringkat
- Livre D'anatomie - KaminaDokumen6 halamanLivre D'anatomie - KaminaFinoana arivony RasolofondrainibeBelum ada peringkat
- L2 CM1 AnatomieDokumen39 halamanL2 CM1 AnatomiejhonronBelum ada peringkat
- RX EpauleDokumen46 halamanRX EpauleMessaoud DehinaBelum ada peringkat
- Thorax-2 0Dokumen67 halamanThorax-2 0Hichem100% (2)
- L'anatomie HumaineDokumen195 halamanL'anatomie Humaineralucag_1100% (2)
- Excitabilite Cellulaire Et Activites Electriques de La CelluleDokumen7 halamanExcitabilite Cellulaire Et Activites Electriques de La CelluleSylvain SalifouBelum ada peringkat
- Effet Tunnel - 8heDokumen7 halamanEffet Tunnel - 8heHicham Al FalouBelum ada peringkat
- Potentiel de Repos Et D'actionDokumen28 halamanPotentiel de Repos Et D'actionWahed Tefla0% (1)
- Le Metabolisme Energetique DR GuedjatiDokumen20 halamanLe Metabolisme Energetique DR GuedjatiMedMan689% (9)
- La Physiologie Du Systeme Nerveux Centrale DR GuedjatiDokumen33 halamanLa Physiologie Du Systeme Nerveux Centrale DR GuedjatiMedMan6100% (2)
- La Physiologie Renale DR GuedjatiDokumen23 halamanLa Physiologie Renale DR GuedjatiMedMan6100% (5)
- La Physiologie Du Milieu Interieur DR GuedjatiDokumen11 halamanLa Physiologie Du Milieu Interieur DR GuedjatiMedMan690% (10)
- La Physiologie Du Systeme Nerveux Autonome DR GuedjatiDokumen16 halamanLa Physiologie Du Systeme Nerveux Autonome DR GuedjatiMedMan675% (4)
- La Physiologie de L Appareil Respiratoire DR GuedjatiDokumen30 halamanLa Physiologie de L Appareil Respiratoire DR GuedjatiMedMan685% (13)
- La Physiologie de La Digestion DR GuedjatiDokumen23 halamanLa Physiologie de La Digestion DR GuedjatiMedMan686% (7)
- Le Milieu Intrieur (Équilibre Hydro-Sodé) 1'Dokumen7 halamanLe Milieu Intrieur (Équilibre Hydro-Sodé) 1'MedMan6100% (1)
- L Exploration Fonctionnelle Renale DR GuedjatiDokumen14 halamanL Exploration Fonctionnelle Renale DR GuedjatiMedMan6100% (12)
- La Physiologie Cardiocirculatoire DR GuedjatiDokumen35 halamanLa Physiologie Cardiocirculatoire DR GuedjatiMedMan6100% (2)
- Le Milieu Interieur (Equilibre Hydro-Sodé) 1Dokumen13 halamanLe Milieu Interieur (Equilibre Hydro-Sodé) 1MedMan6100% (2)
- Le Milieu InterieurDokumen30 halamanLe Milieu InterieurMedMan6100% (9)
- Bioenergie 2Dokumen31 halamanBioenergie 2MedMan6100% (4)
- I Fire ModelisationDokumen71 halamanI Fire Modelisationismailines100% (1)
- Chapitre 5 Polarisation de La Lumière Rectiligne Et RotatoireDokumen2 halamanChapitre 5 Polarisation de La Lumière Rectiligne Et RotatoireBeatrice Florin100% (1)
- Resolution Equation DifferentielleDokumen6 halamanResolution Equation DifferentielletortorouBelum ada peringkat
- Analyse Fonctionnelle:: Energie Électrique Energie MécaniqueDokumen22 halamanAnalyse Fonctionnelle:: Energie Électrique Energie MécaniqueÂÿâčhê Îsmâïl0% (1)
- C03 CoursDokumen4 halamanC03 Coursc_landelBelum ada peringkat
- Série 2 2020-21Dokumen2 halamanSérie 2 2020-21halima rajiBelum ada peringkat
- Poly Af m1Dokumen117 halamanPoly Af m1Laid MerabtiBelum ada peringkat
- Cours Physique 04Dokumen3 halamanCours Physique 04Hanine HmidaBelum ada peringkat
- 1314 Electromagnétisme 2Dokumen6 halaman1314 Electromagnétisme 2Anas HasniBelum ada peringkat
- CdiffCapes PDFDokumen2 halamanCdiffCapes PDFTko ToolBelum ada peringkat
- Chingatome-Troisième-La ProportionnalitéDokumen4 halamanChingatome-Troisième-La ProportionnalitéFouzia BadiBelum ada peringkat
- Ch4 La Traction SimpleDokumen5 halamanCh4 La Traction SimpleMouhi MouhaBelum ada peringkat
- CBP AGAP GRA01 Rev01 Microgravimetrie RFDokumen5 halamanCBP AGAP GRA01 Rev01 Microgravimetrie RFNajib EL GOUMIBelum ada peringkat
- 1 1 ConformesDokumen2 halaman1 1 Conformesckhaled3Belum ada peringkat
- Exos Corr PDFDokumen12 halamanExos Corr PDFryadh100% (1)
- Réduction Des EndomorphismesDokumen70 halamanRéduction Des EndomorphismesIlassa Savadogo100% (1)
- La Variable Aleatoire ContinueDokumen18 halamanLa Variable Aleatoire ContinueBMA-medecine100% (2)
- Résume Mécanique Des FluidesDokumen9 halamanRésume Mécanique Des Fluidessimo100% (4)
- CoursTM1 PDFDokumen60 halamanCoursTM1 PDFMohamed HachimiBelum ada peringkat
- La Contingence: Le Maillon Entre L'irréversibilité Du Temps Et Le MalDokumen78 halamanLa Contingence: Le Maillon Entre L'irréversibilité Du Temps Et Le MalASFDPBelum ada peringkat
- 13813Dokumen5 halaman13813akreitBelum ada peringkat
- Quant Matie Rex oDokumen3 halamanQuant Matie Rex oAbdelouahed OuarrarBelum ada peringkat
- Isostatisme - Mise en Position - Maintien en Position - MIP - MAPDokumen5 halamanIsostatisme - Mise en Position - Maintien en Position - MIP - MAPSamir2020100% (1)
- Courant ContinuDokumen111 halamanCourant ContinubenkhelilBelum ada peringkat
- Rapport 2Dokumen7 halamanRapport 2elking gingstarsBelum ada peringkat