Anda mungkin juga menyukai
- Coagulation Adsorption Ultrafiltration For Wastewater Treatment and Reuse - 2000 - DesalinationDokumen8 halamanCoagulation Adsorption Ultrafiltration For Wastewater Treatment and Reuse - 2000 - DesalinationAlvaro HueteBelum ada peringkat
- Anaerobic Ammonium Oxidation (Anammox) in The Marine EnvironmentDokumen8 halamanAnaerobic Ammonium Oxidation (Anammox) in The Marine EnvironmentKirana agist wangsa putriBelum ada peringkat
- Use of Crossflow Microfiltration in Wastewater Treatment - 1997 - Water ResearchDokumen9 halamanUse of Crossflow Microfiltration in Wastewater Treatment - 1997 - Water ResearchAlvaro HueteBelum ada peringkat
- Wastewater Treatment Performance and Sludge Reduction Properties of A Household Wastewater Treatment System Combined With An Aerobic Sludge DigestionDokumen7 halamanWastewater Treatment Performance and Sludge Reduction Properties of A Household Wastewater Treatment System Combined With An Aerobic Sludge DigestionAlvaro HueteBelum ada peringkat
- Ammonium Removal From Concentrated Waste Streams With The Anaerobic Ammonium Oxidation (Anammox) Process in Different Reactor ConfigurationsDokumen8 halamanAmmonium Removal From Concentrated Waste Streams With The Anaerobic Ammonium Oxidation (Anammox) Process in Different Reactor ConfigurationsAlvaro HueteBelum ada peringkat
- Anaerobic Ammonium Oxidation by Nitrite Anammox Implications For N2 Production in Coastal Marine Sediments - 2005 - Geochimica Et Cosmochimica ActaDokumen9 halamanAnaerobic Ammonium Oxidation by Nitrite Anammox Implications For N2 Production in Coastal Marine Sediments - 2005 - Geochimica Et Cosmochimica ActaAlvaro HueteBelum ada peringkat
- Treatment of The Textile Wastewater by Combined Electrocoagulation - 2006 - ChemosphereDokumen7 halamanTreatment of The Textile Wastewater by Combined Electrocoagulation - 2006 - ChemosphereAlvaro HueteBelum ada peringkat
- The SHARON®-Anammox® Process For Treatment of Ammonium Rich WastewaterDokumen8 halamanThe SHARON®-Anammox® Process For Treatment of Ammonium Rich WastewaterAlvaro HueteBelum ada peringkat
- MBR - SBRDokumen11 halamanMBR - SBRAlvaro HueteBelum ada peringkat
- MBR + AluminioDokumen23 halamanMBR + AluminioAlvaro HueteBelum ada peringkat
- Prueba de Filtración MBRDokumen10 halamanPrueba de Filtración MBRAlvaro HueteBelum ada peringkat
- Completely Autotrophic Nitrogen Removal Over Nitrite in One Single Reactor - 2002 - Water ResearchDokumen8 halamanCompletely Autotrophic Nitrogen Removal Over Nitrite in One Single Reactor - 2002 - Water ResearchAlvaro HueteBelum ada peringkat
- Waste ManagementDokumen8 halamanWaste ManagementAlvaro HueteBelum ada peringkat
- Control of Organic Loading Rate Using The Specific Methanogenic Activity Test During Start-Up of A N Anaerobic Digestion SystemDokumen7 halamanControl of Organic Loading Rate Using The Specific Methanogenic Activity Test During Start-Up of A N Anaerobic Digestion SystemAlvaro HueteBelum ada peringkat
- Minor Thesis G.Schutgens PDFDokumen66 halamanMinor Thesis G.Schutgens PDFAlvaro HueteBelum ada peringkat
- Minor Thesis G.Schutgens PDFDokumen66 halamanMinor Thesis G.Schutgens PDFAlvaro HueteBelum ada peringkat
- Anaerobic Sludge Blanket ReactorsDokumen17 halamanAnaerobic Sludge Blanket ReactorsAlvaro HueteBelum ada peringkat
- Monitoring of Specific Methanogenic Activity of Granular Sludge by Confocal Laser Scanning Microscopy During Start-Up of Thermophilic Upflow Anaerobic Sludge Blanket ReactorDokumen6 halamanMonitoring of Specific Methanogenic Activity of Granular Sludge by Confocal Laser Scanning Microscopy During Start-Up of Thermophilic Upflow Anaerobic Sludge Blanket ReactorAlvaro HueteBelum ada peringkat
- Granulation For Coking Wastewater Treatment in A CDokumen6 halamanGranulation For Coking Wastewater Treatment in A CAlvaro HueteBelum ada peringkat
- Inputoutput Linearization Control Technique For AnDokumen6 halamanInputoutput Linearization Control Technique For AnAlvaro HueteBelum ada peringkat
- Meteor - MBBR - P ER 011 EN 1512 PDFDokumen2 halamanMeteor - MBBR - P ER 011 EN 1512 PDFAlvaro HueteBelum ada peringkat
- Biomod: Packaged Moving Bed Biofilm ReactorDokumen4 halamanBiomod: Packaged Moving Bed Biofilm ReactorAlvaro Huete100% (1)
- Applying Internal Circulation Anaerobic Reactor FoDokumen8 halamanApplying Internal Circulation Anaerobic Reactor FoAlvaro HueteBelum ada peringkat
- Wastewater IndustrialDokumen12 halamanWastewater IndustrialAlvaro HueteBelum ada peringkat
- Air LiftpresentationDokumen15 halamanAir LiftpresentationTrushit MakwanaBelum ada peringkat
- IndWWTreat ActSld Isbn1843391449 ContentsDokumen9 halamanIndWWTreat ActSld Isbn1843391449 ContentsAlvaro HueteBelum ada peringkat
- Simulation and Calibration of A Full-Scale SequencDokumen10 halamanSimulation and Calibration of A Full-Scale SequencAlvaro HueteBelum ada peringkat
- Filtro PercoladorDokumen8 halamanFiltro PercoladorAlvaro HueteBelum ada peringkat
- Wastewater IndustrialDokumen12 halamanWastewater IndustrialAlvaro HueteBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Analysis and Design of Well FoundationDokumen40 halamanAnalysis and Design of Well Foundationdhanabal100% (1)
- Assistant Bookkeeper Resume Sample - Best Format - Great Sample ResumeDokumen4 halamanAssistant Bookkeeper Resume Sample - Best Format - Great Sample ResumedrustagiBelum ada peringkat
- High Performance Vector Control SE2 Series InverterDokumen9 halamanHigh Performance Vector Control SE2 Series InverterhanazahrBelum ada peringkat
- Integration Plan Grade 9 Mapeh SeptemberDokumen3 halamanIntegration Plan Grade 9 Mapeh Septemberbernie evaristo bacsaBelum ada peringkat
- Thesis StoryboardDokumen5 halamanThesis StoryboardJill Brown100% (2)
- INSTRUCTIONAL SUPERVISORY PLAN 1st Quarter of SY 2023 2024 Quezon ISDokumen7 halamanINSTRUCTIONAL SUPERVISORY PLAN 1st Quarter of SY 2023 2024 Quezon ISayongaogracelyflorBelum ada peringkat
- Introduction To The Iceberg ModelDokumen5 halamanIntroduction To The Iceberg ModelAbhay Tiwari100% (1)
- ER288 090714 5082 CV OKP (089) Method Statement For Plate Baring TestDokumen3 halamanER288 090714 5082 CV OKP (089) Method Statement For Plate Baring TestWr ArBelum ada peringkat
- Tools of Persuasion StudentsDokumen4 halamanTools of Persuasion StudentsBelén Revilla GonzálesBelum ada peringkat
- 2SA1016Dokumen4 halaman2SA1016catalina maryBelum ada peringkat
- Focal Length of Convex LensDokumen5 halamanFocal Length of Convex LensHey AnuragBelum ada peringkat
- Excavation Trench Permit Ex 1 F0206Dokumen5 halamanExcavation Trench Permit Ex 1 F0206emeka2012Belum ada peringkat
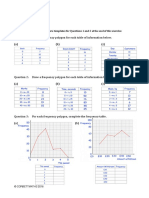
- Frequency Polygons pdf3Dokumen7 halamanFrequency Polygons pdf3Nevine El shendidyBelum ada peringkat
- S010T1Dokumen1 halamanS010T1DUCBelum ada peringkat
- Dady - Piernas LargasDokumen12 halamanDady - Piernas LargasSarha NietoBelum ada peringkat
- Slip Ring Motor CheckDokumen3 halamanSlip Ring Motor CheckRolando LoayzaBelum ada peringkat
- STFC-2023 International E - conference-BITDokumen6 halamanSTFC-2023 International E - conference-BITRanilprabhu MBelum ada peringkat
- Hunger Games Mini Socratic Seminar2012Dokumen4 halamanHunger Games Mini Socratic Seminar2012Cary L. TylerBelum ada peringkat
- Riveted JointsDokumen28 halamanRiveted Jointsgnbabuiitg0% (1)
- Eurocode Conference 2023 Seters Van Eurocode 7Dokumen33 halamanEurocode Conference 2023 Seters Van Eurocode 7Rodolfo BlanchiettiBelum ada peringkat
- Harmony Guide DatabaseDokumen7 halamanHarmony Guide DatabaseAya SakamotoBelum ada peringkat
- AB-005-2020 Dated 10.09.2020 (SKF-Prestine)Dokumen3 halamanAB-005-2020 Dated 10.09.2020 (SKF-Prestine)AliasgarBelum ada peringkat
- IVISOR Mentor IVISOR Mentor QVGADokumen2 halamanIVISOR Mentor IVISOR Mentor QVGAwoulkanBelum ada peringkat
- R67068.0002 2 HB Profibus-Schnittstelle en KueblerDokumen42 halamanR67068.0002 2 HB Profibus-Schnittstelle en KueblerSabari StunnerBelum ada peringkat
- Postmodernism in Aha! MovieDokumen2 halamanPostmodernism in Aha! MovieSayma AkterBelum ada peringkat
- Contoh Label Sensus 2022Dokumen313 halamanContoh Label Sensus 2022Ajenk SablackBelum ada peringkat
- On The Importance of Learning Statistics For Psychology StudentsDokumen2 halamanOn The Importance of Learning Statistics For Psychology StudentsMadison HartfieldBelum ada peringkat
- Bearing TypesDokumen5 halamanBearing TypesWayuBelum ada peringkat
- WallthicknessDokumen1 halamanWallthicknessGabriela MotaBelum ada peringkat
- Case Study III - MichelinDokumen15 halamanCase Study III - MichelinfreitzBelum ada peringkat