Anda mungkin juga menyukai
- Micro HCTDokumen1 halamanMicro HCTIgor MagalhãesBelum ada peringkat
- NR CurveDokumen1 halamanNR CurvePhúc Bùi VănBelum ada peringkat
- MS Word 4 (Insert/chart ) : Bat Dulam Suren DorjDokumen1 halamanMS Word 4 (Insert/chart ) : Bat Dulam Suren DorjNaraa JaminBelum ada peringkat
- 2 MTPH Milling Section (RAVA)Dokumen1 halaman2 MTPH Milling Section (RAVA)Pranav GoyalBelum ada peringkat
- Tamiz PASA % Arena AnalisisDokumen24 halamanTamiz PASA % Arena AnalisisElvis Armando Huaman HuamaniBelum ada peringkat
- MIC - Project-Group-04.vi E:/MIC Lab/MIC - Project-Group-04.vi Last Modified On 22/03/21 at 2:59 PM Printed On 22/03/21 at 3:31 PMDokumen1 halamanMIC - Project-Group-04.vi E:/MIC Lab/MIC - Project-Group-04.vi Last Modified On 22/03/21 at 2:59 PM Printed On 22/03/21 at 3:31 PMMd. Ar-Rafi Bin Arif 190011123Belum ada peringkat
- Comple, Soares, 2019Dokumen11 halamanComple, Soares, 2019SOBelum ada peringkat
- PETA TOPOGRAFI BABODokumen1 halamanPETA TOPOGRAFI BABOM taufik AshariBelum ada peringkat
- 5G Bandwise Throught TrendDokumen6 halaman5G Bandwise Throught TrendabhineetkumarBelum ada peringkat
- 123Dokumen1 halaman123Harish Kumar.THOTABelum ada peringkat
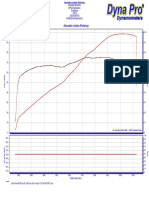
- Adrenaline Junkies Workshop: 3-Dyno Run - DPR, Dyno Run, 17/05/2019, 18:43:26Dokumen1 halamanAdrenaline Junkies Workshop: 3-Dyno Run - DPR, Dyno Run, 17/05/2019, 18:43:26Βασίλης ΝαλμπάντηςBelum ada peringkat
- ASEREP v5.2.0.0Dokumen1 halamanASEREP v5.2.0.0Ali EscobarBelum ada peringkat
- Tracer USGSDokumen72 halamanTracer USGSAdriana RaciniBelum ada peringkat
- Historicdata - Ping - 2021 06 16 00 00 00 - 2021 06 17 00 00 00Dokumen2 halamanHistoricdata - Ping - 2021 06 16 00 00 00 - 2021 06 17 00 00 00ashishladooBelum ada peringkat
- Allan DroopyDokumen1 halamanAllan DroopyLuis MiddletonBelum ada peringkat
- Catalog of quick-release hydraulic couplingsDokumen404 halamanCatalog of quick-release hydraulic couplingsmecelberBelum ada peringkat
- F5010 FM and TV2-6 Blue CurvesDokumen1 halamanF5010 FM and TV2-6 Blue Curvescostello202Belum ada peringkat
- Blood Sampling GuideDokumen4 halamanBlood Sampling GuideJosue GarciaBelum ada peringkat
- Allan LarvaDokumen1 halamanAllan LarvaLuis MiddletonBelum ada peringkat
- Allan Martin SeptumDokumen1 halamanAllan Martin SeptumLuis MiddletonBelum ada peringkat
- Exalted2 Character Sheet DragonBlooded v1.1Dokumen2 halamanExalted2 Character Sheet DragonBlooded v1.1Lenice EscadaBelum ada peringkat
- NATAL TUBA BBDokumen1 halamanNATAL TUBA BBCentral RDBelum ada peringkat
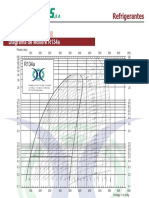
- Diagrama de Molière R134a: RefrigerantesDokumen1 halamanDiagrama de Molière R134a: RefrigerantesHumnerBelum ada peringkat
- Tauane Raizila Ferreira Dos Santos-Tauane Raizila Ferreira Dos Santos MX 3Dokumen5 halamanTauane Raizila Ferreira Dos Santos-Tauane Raizila Ferreira Dos Santos MX 3tauaneraizila15Belum ada peringkat
- Fvo Panel - Schematic Latest Updated 18.10.2023Dokumen18 halamanFvo Panel - Schematic Latest Updated 18.10.2023kirubahnanthanBelum ada peringkat
- Allan Dewey 40 Canadiense Arizonaa: Sacerdote / Ex Bandido MasculinoDokumen1 halamanAllan Dewey 40 Canadiense Arizonaa: Sacerdote / Ex Bandido MasculinoLuis MiddletonBelum ada peringkat
- Allan Martin OhmydogDokumen1 halamanAllan Martin OhmydogLuis MiddletonBelum ada peringkat
- Allan Martin SepterDokumen1 halamanAllan Martin SepterLuis MiddletonBelum ada peringkat
- Book 1Dokumen2 halamanBook 1paapuhomeBelum ada peringkat
- Grain Size Distribution AnalysisDokumen1 halamanGrain Size Distribution AnalysisDileep K NambiarBelum ada peringkat
- % Ter Tiar y Industr: For Examiner's UseDokumen2 halaman% Ter Tiar y Industr: For Examiner's UseRana SlimBelum ada peringkat
- Corpiño ImprimirDokumen1 halamanCorpiño ImprimirSebastianBelum ada peringkat
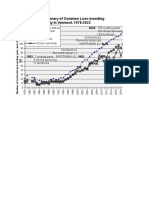
- Summary of Common Loon Breeding Activity From 1978-2022, Courtesy of Eric Hanson and Vermont Center For Ecostudies.Dokumen2 halamanSummary of Common Loon Breeding Activity From 1978-2022, Courtesy of Eric Hanson and Vermont Center For Ecostudies.Kate SadoffBelum ada peringkat
- Kurva Washability Test FixDokumen2 halamanKurva Washability Test FixWita ManaluBelum ada peringkat
- Grain Size Distribution Graph Aggregate GradationDokumen1 halamanGrain Size Distribution Graph Aggregate GradationchrishoppepeBelum ada peringkat
- KBL Pump GADokumen11 halamanKBL Pump GAudgeetBelum ada peringkat
- Character Sheet: Hero Attributes and SkillsDokumen1 halamanCharacter Sheet: Hero Attributes and SkillsRobert David KeenBelum ada peringkat
- Cooling Capacity 3 Tons - Temperature Difference: CASE 2 (Homework) (5 Ptos)Dokumen4 halamanCooling Capacity 3 Tons - Temperature Difference: CASE 2 (Homework) (5 Ptos)Ronaldo CTBelum ada peringkat
- Test Administrations: 01/03/20, 04/05/20, 08/01/20: Progress Report ID: 12345 SCL-90-R Norms: Adult NonpatientDokumen2 halamanTest Administrations: 01/03/20, 04/05/20, 08/01/20: Progress Report ID: 12345 SCL-90-R Norms: Adult NonpatientRim InfBelum ada peringkat
- PDF - PdpiefDokumen4 halamanPDF - Pdpiefhowa howaBelum ada peringkat
- 1 - Southern Cross - Iso Pump - 50 X 32 - 160 - 1410-2880 RPM PDFDokumen1 halaman1 - Southern Cross - Iso Pump - 50 X 32 - 160 - 1410-2880 RPM PDFIra LtrBelum ada peringkat
- Pareto Charts Explained for Identifying Main Causes of ProblemsDokumen5 halamanPareto Charts Explained for Identifying Main Causes of ProblemsTrexie PalenciaBelum ada peringkat
- Grafi k Nilai pre test dan post testDokumen1 halamanGrafi k Nilai pre test dan post test20-107 DIMAS AGMA MAHENDRABelum ada peringkat
- Sieve Analysis GraphDokumen1 halamanSieve Analysis GraphEdo VandimaBelum ada peringkat
- Merry Go Round-ViolínDokumen1 halamanMerry Go Round-ViolínCecilia AnchondoBelum ada peringkat
- Devaki ItwsDokumen3 halamanDevaki Itwsswarna_793238588Belum ada peringkat
- MARRY YOU ViolaDokumen1 halamanMARRY YOU ViolaHERNANDARIO HERNANDEZBelum ada peringkat
- Piping Load VapourDokumen1 halamanPiping Load VapourSơn Nguyễn-LêBelum ada peringkat
- X Bar Chart: Machine NameDokumen2 halamanX Bar Chart: Machine NameVIBHORBelum ada peringkat
- PRAKBIOFAR - C - Orin Chia Elga - 08061381823063 - 5Dokumen2 halamanPRAKBIOFAR - C - Orin Chia Elga - 08061381823063 - 5Orien Chia ElgaBelum ada peringkat
- Kerikil Kecil 10 MM - 15 MM Kerikil BesarDokumen1 halamanKerikil Kecil 10 MM - 15 MM Kerikil BesarMaulid Abdullah LuthfirrachmanBelum ada peringkat
- Manpower-Histogram - Crude Oil Tank - 20-May-2017-1Dokumen1 halamanManpower-Histogram - Crude Oil Tank - 20-May-2017-1sunjeyBelum ada peringkat
- PR OV ED: A Graphical Representation of Inverse VRML UptakeDokumen1 halamanPR OV ED: A Graphical Representation of Inverse VRML UptakeMahbubRiyadBelum ada peringkat
- Grain Size Distribution: Client Mbits Project Name P05 Project Number Po5Dokumen1 halamanGrain Size Distribution: Client Mbits Project Name P05 Project Number Po5Dileep K NambiarBelum ada peringkat
- Article I GraphDokumen1 halamanArticle I GraphKumar SingamBelum ada peringkat
- Absensi Konseling Gizi (Respons)Dokumen2 halamanAbsensi Konseling Gizi (Respons)Muhammad RizalBelum ada peringkat
- Modul Anu FRNTDokumen1 halamanModul Anu FRNTMuhammad RizalBelum ada peringkat
- Audiensi Menkes RIDokumen1 halamanAudiensi Menkes RIMuhammad RizalBelum ada peringkat
- A SalvadorDokumen1 halamanA SalvadorMuhammad RizalBelum ada peringkat
- Introduction To Public Health Nutrition: DR Nayyar Raza Kazmi MBBS, DHPM, MPH, M.SCDokumen11 halamanIntroduction To Public Health Nutrition: DR Nayyar Raza Kazmi MBBS, DHPM, MPH, M.SCMuhammad RizalBelum ada peringkat
- Transcript Trump's Speech About PalestinaDokumen14 halamanTranscript Trump's Speech About PalestinaMuhammad RizalBelum ada peringkat
- Effort of Treatment and Behavior of Diabetes Patients in MakassarDokumen1 halamanEffort of Treatment and Behavior of Diabetes Patients in MakassarMuhammad RizalBelum ada peringkat
- Pragmatics and StylisticsDokumen7 halamanPragmatics and StylisticsAri A. ResulBelum ada peringkat
- Edit Pra FinalDokumen47 halamanEdit Pra FinalMuhammad RizalBelum ada peringkat
- DsfggwsfgwsfwsDokumen1 halamanDsfggwsfgwsfwsMuhammad RizalBelum ada peringkat
- Brochure FinalDokumen14 halamanBrochure FinalRajesh BadheBelum ada peringkat
- Re Insurance BrochureDokumen12 halamanRe Insurance BrochureQBE European OperationsBelum ada peringkat
- Categories of AnalysisDokumen4 halamanCategories of AnalysisTee JackBelum ada peringkat
- Elastosil® M 4642 A/B: Product DescriptionDokumen2 halamanElastosil® M 4642 A/B: Product DescriptionhemantrulzBelum ada peringkat
- LPT22Dokumen3 halamanLPT22Leonardo Vinicio Olarte CarrilloBelum ada peringkat
- Child Development: Resilience & Risk, A Course SyllabusDokumen17 halamanChild Development: Resilience & Risk, A Course SyllabusJane GilgunBelum ada peringkat
- Endangered Species Lesson PlanDokumen5 halamanEndangered Species Lesson PlanIrena Stamenova100% (1)
- Ficha Tecnica Sonometro Artisan SL2100PSDokumen1 halamanFicha Tecnica Sonometro Artisan SL2100PSJackson Steward Ríos SastoqueBelum ada peringkat
- Extent of Compliance on Waste Disposal PracticesDokumen13 halamanExtent of Compliance on Waste Disposal PracticesEzza Jean Paypa100% (3)
- Bladder StonesDokumen20 halamanBladder StonesRiean AuliaBelum ada peringkat
- Attention Deficit Hyperactivity Disorder (ADHD) in ChildrenDokumen16 halamanAttention Deficit Hyperactivity Disorder (ADHD) in ChildrenVenmari Joyce Pasinos100% (1)
- PH 611: CO2 Release in RespirationDokumen2 halamanPH 611: CO2 Release in RespirationAcuCJamBelum ada peringkat
- Apd440c 6Dokumen5 halamanApd440c 6Cristhian ChoqueBelum ada peringkat
- Sensation As If by Roberts PDFDokumen369 halamanSensation As If by Roberts PDFNauman Khan100% (1)
- The Liver in Systemic Disease A Clinician's Guide To Abnormal LiverDokumen285 halamanThe Liver in Systemic Disease A Clinician's Guide To Abnormal Liveroleksandra.bilotkachBelum ada peringkat
- Manual Reparatii Audi A2 1.6 FSI PDFDokumen104 halamanManual Reparatii Audi A2 1.6 FSI PDFferentz u100% (1)
- Homework 1Dokumen10 halamanHomework 1Nam Anh KiraiBelum ada peringkat
- Transplantation ActDokumen24 halamanTransplantation ActHomer MoeBelum ada peringkat
- Pehealth11 q2 Mod2of2 H.O.P.E v2Dokumen26 halamanPehealth11 q2 Mod2of2 H.O.P.E v2Avillz Mar LeeBelum ada peringkat
- Sustainable City PDFDokumen361 halamanSustainable City PDFagusbudiyonoBelum ada peringkat
- Online Shopping ProjectDokumen8 halamanOnline Shopping Projectsrilakshmi itham100% (1)
- KeirseyDokumen28 halamanKeirseyapi-525703700Belum ada peringkat
- Medical Student Amnesia USMLE Step 1 - Flash Cards by CueFlashDokumen13 halamanMedical Student Amnesia USMLE Step 1 - Flash Cards by CueFlashMuhammad Farhan KhaliqBelum ada peringkat
- Panduan Sejarah Ekologi Taman Nasional Komodo Buku 2Dokumen228 halamanPanduan Sejarah Ekologi Taman Nasional Komodo Buku 2Agi SemBelum ada peringkat
- Rabor vs CSC - Retirement age extension disputeDokumen2 halamanRabor vs CSC - Retirement age extension disputeGladys Bustria OrlinoBelum ada peringkat
- UntitledDokumen10 halamanUntitledGuia Charish SalvadorBelum ada peringkat
- The Political Ecology of Food and AgricultureDokumen15 halamanThe Political Ecology of Food and AgricultureMitchBelum ada peringkat
- Doble M4000 User GuideDokumen204 halamanDoble M4000 User GuidePablo Toro Lopez100% (1)
- 3G3EV Installation ManualDokumen55 halaman3G3EV Installation ManualHajrudin SinanovićBelum ada peringkat
- Effects of Air and Noise PollutionDokumen6 halamanEffects of Air and Noise Pollutionhaiqa malikBelum ada peringkat