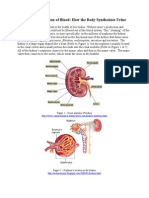
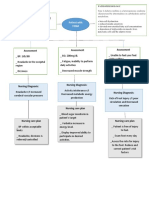
Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- BPH, Pdui 2017 PDFDokumen36 halamanBPH, Pdui 2017 PDFcyndra erisBelum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- EcDokumen1 halamanEcMega Redha PutriBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- 0-2707558646 20210611 090951 0000Dokumen9 halaman0-2707558646 20210611 090951 0000Mega Redha PutriBelum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Juli 2017Dokumen2 halamanJuli 2017Mega Redha PutriBelum ada peringkat
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Jadwal Tampil Kelompok I: (BST, Ronde) (BST) (BST)Dokumen1 halamanJadwal Tampil Kelompok I: (BST, Ronde) (BST) (BST)Mega Redha PutriBelum ada peringkat
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- A Chicago Tradition Since 1949: Citylink Mall Resorts World SentosaDokumen1 halamanA Chicago Tradition Since 1949: Citylink Mall Resorts World SentosaMega Redha PutriBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Scholarship Guide English 20150721Dokumen6 halamanScholarship Guide English 20150721M Muchdif Al AfghoniBelum ada peringkat
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- BPH, Pdui 2017 PDFDokumen36 halamanBPH, Pdui 2017 PDFcyndra erisBelum ada peringkat
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Big Little LiesDokumen1 halamanBig Little LiesMega Redha Putri100% (1)
- Jurnal Varisela InggriDokumen3 halamanJurnal Varisela InggriMega Redha PutriBelum ada peringkat
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- EcDokumen1 halamanEcMega Redha PutriBelum ada peringkat
- Surat Izin ElektraDokumen1 halamanSurat Izin ElektraMega Redha PutriBelum ada peringkat
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Members Name in HangulDokumen1 halamanMembers Name in HangulMega Redha PutriBelum ada peringkat
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Roman IzationDokumen3 halamanRoman IzationMega Redha PutriBelum ada peringkat
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- 6th Central Pay Commission Salary CalculatorDokumen15 halaman6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Grad SpeechDokumen1 halamanGrad SpeechMega Redha PutriBelum ada peringkat
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Auto TextDokumen2 halamanAuto TextMega Redha PutriBelum ada peringkat
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- Lake TobaDokumen1 halamanLake TobaMega Redha PutriBelum ada peringkat
- Propofol Anesthesia EffectsDokumen4 halamanPropofol Anesthesia EffectsdidyahfBelum ada peringkat
- A Comprehensive Review of Global Efforts on COVID-19 Vaccine DevelopmentDokumen22 halamanA Comprehensive Review of Global Efforts on COVID-19 Vaccine DevelopmentBeverly EstoqueBelum ada peringkat
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Bone Grafting: Classification by OriginDokumen4 halamanBone Grafting: Classification by OriginHerlangga Fadhillah AkbarBelum ada peringkat
- Anxiety DisordersDokumen20 halamanAnxiety DisordersAngela BautistaBelum ada peringkat
- Lab Manual GeneticsDokumen15 halamanLab Manual Geneticsapi-319985329100% (1)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- CoenzymesDokumen60 halamanCoenzymesraddagBelum ada peringkat
- DILSHAD - Study DesignsDokumen48 halamanDILSHAD - Study DesignsAamir IjazBelum ada peringkat
- Simina Maria Boca, PH.D.: EducationDokumen13 halamanSimina Maria Boca, PH.D.: EducationRohit AroraBelum ada peringkat
- Handbook of Pediatric Hematology and Oncology 3rd Edition 2021Dokumen492 halamanHandbook of Pediatric Hematology and Oncology 3rd Edition 2021pgginecologiahospirioBelum ada peringkat
- 3rd Quarter Exam Science 10Dokumen3 halaman3rd Quarter Exam Science 10Rizel Shaira Hope TanamanBelum ada peringkat
- Conceptual Frameworks and Theories in Psychiatric - Mental Health NursingDokumen124 halamanConceptual Frameworks and Theories in Psychiatric - Mental Health NursingHui Chen100% (1)
- HYbripol DNA PolymeraseDokumen1 halamanHYbripol DNA PolymeraseDanuBelum ada peringkat
- Borrador Writing 3Dokumen3 halamanBorrador Writing 3Carlos LazoBelum ada peringkat
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- 2402 CH 17 Endocrine System (Part 1) PDFDokumen23 halaman2402 CH 17 Endocrine System (Part 1) PDFHarry RussellBelum ada peringkat
- Biomedical Research To Improve The Lives of People With Down SyndromeDokumen13 halamanBiomedical Research To Improve The Lives of People With Down SyndromeGlobalDownSyndromeBelum ada peringkat
- NephronDokumen5 halamanNephronrdeguzma100% (1)
- Inheritance of Sex and Sex-Linked or Influenced Traits - StudentDokumen21 halamanInheritance of Sex and Sex-Linked or Influenced Traits - StudentNoob Gamer0% (1)
- MG PosterDokumen1 halamanMG PosterNataShini RajaRatnamBelum ada peringkat
- Concept MappingDokumen2 halamanConcept MappingNhindoi KonhinnoiBelum ada peringkat
- Physiology and Neurobiology of Stress and Adaptation - Central Role of The BrainDokumen33 halamanPhysiology and Neurobiology of Stress and Adaptation - Central Role of The BrainJSGIBelum ada peringkat
- 3 CellsDokumen7 halaman3 CellsCaviles, Jasmin S.Belum ada peringkat
- VPA & Glia &TLR Deckmann-2018-Autism-ADokumen15 halamanVPA & Glia &TLR Deckmann-2018-Autism-AAhmed NagehBelum ada peringkat
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Eucerin - Nobel Prize Revolutionises Skin Care - Cells' Water Channels Specifically StimulatedDokumen2 halamanEucerin - Nobel Prize Revolutionises Skin Care - Cells' Water Channels Specifically StimulatedFelpnilBelum ada peringkat
- Antigen Processing and Presentation 09Dokumen34 halamanAntigen Processing and Presentation 09Khairul Ikhwan100% (2)
- Mezoterapija PDFDokumen11 halamanMezoterapija PDFDavid DijakBelum ada peringkat
- The Enigma of Death ExplainedDokumen19 halamanThe Enigma of Death Explainedsorrisodevoltaire0% (1)
- 5t4bvrfke3xuqspihwrir0d1Dokumen6 halaman5t4bvrfke3xuqspihwrir0d1Vishesh Mahendru20% (5)
- Jessica Hughes AP Biology LabDokumen8 halamanJessica Hughes AP Biology LabJessica HughesBelum ada peringkat
- Mapping Neural Circuits Using SBEMDokumen7 halamanMapping Neural Circuits Using SBEMIvana JeremicBelum ada peringkat
- Inquisitor Character Creation and AdvancementDokumen10 halamanInquisitor Character Creation and AdvancementMichael MonchampBelum ada peringkat
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDari EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionPenilaian: 4 dari 5 bintang4/5 (402)
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingDari EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingPenilaian: 5 dari 5 bintang5/5 (4)