Anda mungkin juga menyukai
- Introduction To Stem Cell Theraphy Week 8Dokumen10 halamanIntroduction To Stem Cell Theraphy Week 8HappyBelum ada peringkat
- Ideas and Opinions: Promises and Challenges of Stem Cell Research For Regenerative MedicineDokumen9 halamanIdeas and Opinions: Promises and Challenges of Stem Cell Research For Regenerative MedicineKNmanBelum ada peringkat
- What Are Stem Cells and Where Do They Come FromDokumen4 halamanWhat Are Stem Cells and Where Do They Come FromjaniellegbBelum ada peringkat
- Adult Stem CellDokumen13 halamanAdult Stem CellDipti BagdeBelum ada peringkat
- AbstractDokumen7 halamanAbstractLiviu GogataBelum ada peringkat
- Stem Cells in The ClinicDokumen8 halamanStem Cells in The ClinicKamal AdeyemiBelum ada peringkat
- Stem Cell ResearchDokumen4 halamanStem Cell Researchpianomaster82Belum ada peringkat
- (Visvader Et Al., 2016) Tissue-Specific Designs of Stem Cell HierarchiesDokumen7 halaman(Visvader Et Al., 2016) Tissue-Specific Designs of Stem Cell HierarchiesJulio dR AltavasBelum ada peringkat
- Andrews 2002Dokumen14 halamanAndrews 2002Lara P. CarvalhoBelum ada peringkat
- Stem Cell Proposal Final 2003.Dokumen13 halamanStem Cell Proposal Final 2003.Harshal GohilBelum ada peringkat
- The Bone Marrow Niche For Haematopoietic Stem CellsDokumen21 halamanThe Bone Marrow Niche For Haematopoietic Stem CellsHitomi-Belum ada peringkat
- Derivation of Totipotent-Like Stem Cells With Blastocyst-Like Structure Forming PotentialDokumen17 halamanDerivation of Totipotent-Like Stem Cells With Blastocyst-Like Structure Forming PotentialJosué Cristhian Del Valle HornaBelum ada peringkat
- Turning A Skin Into A NeuronDokumen49 halamanTurning A Skin Into A NeuronAngel Daniel Flores AmezcuaBelum ada peringkat
- Adult Stem CellDokumen9 halamanAdult Stem CellMeloveshop DropBelum ada peringkat
- Differences of Stem Cell TypesDokumen12 halamanDifferences of Stem Cell TypesIsmail Tidak Ada KepanjanganBelum ada peringkat
- Adult Stem CellsDokumen9 halamanAdult Stem CellsPedro Henrique100% (1)
- Stem Cells in Regenerative Medicine and Therapeutic PotentialDokumen12 halamanStem Cells in Regenerative Medicine and Therapeutic PotentialIJAR JOURNALBelum ada peringkat
- CMB (Lec)Dokumen94 halamanCMB (Lec)JOHN RONALD BORDOBelum ada peringkat
- Stem Cell: Cellula PrecursoriaDokumen26 halamanStem Cell: Cellula PrecursoriaYhan Brotamonte BoneoBelum ada peringkat
- Stem Cells - A Revolution in Medicine - PDDokumen20 halamanStem Cells - A Revolution in Medicine - PDTatiane Cruz da SilvaBelum ada peringkat
- Stem Cells: Potential for Therapy and Disease InsightsDokumen9 halamanStem Cells: Potential for Therapy and Disease InsightsMahesh T MadhavanBelum ada peringkat
- Gene Therapy and ProspectsDokumen9 halamanGene Therapy and ProspectsAngelBelum ada peringkat
- Farin 2009Dokumen25 halamanFarin 2009Laura Duarte RojasBelum ada peringkat
- Stem Cell TherapyDokumen26 halamanStem Cell TherapymfspongebobBelum ada peringkat
- Chapter 8Dokumen12 halamanChapter 8lala_0Belum ada peringkat
- Cell Theory, Specificity, and Reproduction, 1837-1870: Europe PMC Funders GroupDokumen10 halamanCell Theory, Specificity, and Reproduction, 1837-1870: Europe PMC Funders GroupSilvio SánchezBelum ada peringkat
- Introduction To Stem Cells: Speaker - Maj Mohd Anas Sheikh Guide - LT Col AK BiswasDokumen38 halamanIntroduction To Stem Cells: Speaker - Maj Mohd Anas Sheikh Guide - LT Col AK BiswasMohd Anas SheikhBelum ada peringkat
- 100 Plus Years of Stem Cell Research-20 Years of ISSCRDokumen20 halaman100 Plus Years of Stem Cell Research-20 Years of ISSCRDr Abhigyan ShankarBelum ada peringkat
- Stem CellsDokumen20 halamanStem CellsalibrownBelum ada peringkat
- Your Inner HealersDokumen8 halamanYour Inner HealersAna María TrujilloBelum ada peringkat
- Scaffolds For Stem CellsDokumen8 halamanScaffolds For Stem Cellscubed3Belum ada peringkat
- InTech-Stem Cells Ethical and Religious Issues PDFDokumen17 halamanInTech-Stem Cells Ethical and Religious Issues PDFFionna PohanBelum ada peringkat
- Shamir 2014 - Review organoid 3DDokumen40 halamanShamir 2014 - Review organoid 3DDaniela PatricioBelum ada peringkat
- Stem Cells in Gynecology 2012 American Journal of Obstetrics and GynecologyDokumen8 halamanStem Cells in Gynecology 2012 American Journal of Obstetrics and GynecologyfujimeisterBelum ada peringkat
- Stem Cell: Mihai - Cristian BAGHINA Seria A Grupa 2Dokumen9 halamanStem Cell: Mihai - Cristian BAGHINA Seria A Grupa 2Mihai NycromBelum ada peringkat
- Development of Membrane Like Pre Stem Cells After Released From Tube Shaped NichesDokumen8 halamanDevelopment of Membrane Like Pre Stem Cells After Released From Tube Shaped NichesHerald Scholarly Open AccessBelum ada peringkat
- Signaling Pathways in Cancer and Embryonic Stem CellsDokumen12 halamanSignaling Pathways in Cancer and Embryonic Stem CellsMiftakhul fadliBelum ada peringkat
- Stem Cell: Navigation SearchDokumen14 halamanStem Cell: Navigation Searchanon-589691Belum ada peringkat
- Cell Interactions Guide FateDokumen25 halamanCell Interactions Guide Fatekaladhar reddyBelum ada peringkat
- Stem Cell Research A ReportDokumen7 halamanStem Cell Research A ReportAileen Tan JansorBelum ada peringkat
- Stem Cell ThesisDokumen62 halamanStem Cell ThesisShradha SharmaBelum ada peringkat
- Potential of Induced Pluripotent Stem Cells For Use in Gene TherapyDokumen16 halamanPotential of Induced Pluripotent Stem Cells For Use in Gene TherapyAnnaaaaaBelum ada peringkat
- Regenerative Medicine 2006Dokumen70 halamanRegenerative Medicine 2006subinbioBelum ada peringkat
- Ethical and Legal Issues in Therapeutic CloningDokumen5 halamanEthical and Legal Issues in Therapeutic CloningkazugawaBelum ada peringkat
- (Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Dokumen9 halaman(Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Priscilla FreschuBelum ada peringkat
- Stem CellDokumen9 halamanStem CellDulce UribeBelum ada peringkat
- Stem CellDokumen8 halamanStem CellHanumat SinghBelum ada peringkat
- Mesenchymal Stem CellsDokumen13 halamanMesenchymal Stem CellsArya NugrahaBelum ada peringkat
- Stem Cells Are Unspecialized. One of The Fundamental Properties ofDokumen56 halamanStem Cells Are Unspecialized. One of The Fundamental Properties ofnitishgotBelum ada peringkat
- Embryonic Stem Cells: Review ArticleDokumen6 halamanEmbryonic Stem Cells: Review ArticleFabLab EIUBelum ada peringkat
- Stem CellsDokumen3 halamanStem CellsLhoNa ﭢ ArpilledaBelum ada peringkat
- Understanding Stem CellsDokumen26 halamanUnderstanding Stem CellsNational Academies of Science, Engineering, and Medicine96% (23)
- Key ArticleDokumen9 halamanKey ArticleJosh McHughBelum ada peringkat
- Biology of Stem Cells: An Overview: Pedro C. Chagastelles and Nance B. NardiDokumen5 halamanBiology of Stem Cells: An Overview: Pedro C. Chagastelles and Nance B. NardiMayuri DuttaBelum ada peringkat
- The Momentous Transition To Multicellular Life May Not Have Been So Hard After All - Science - AAASDokumen7 halamanThe Momentous Transition To Multicellular Life May Not Have Been So Hard After All - Science - AAASsathish.botBelum ada peringkat
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsDari EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsPenilaian: 5 dari 5 bintang5/5 (1)
- Diet in Irritable Bowel Syndrome: Review Open AccessDokumen11 halamanDiet in Irritable Bowel Syndrome: Review Open AccessjoseangelroblesBelum ada peringkat
- Manipulation of Gut Microbiota Influences Immune RDokumen18 halamanManipulation of Gut Microbiota Influences Immune RjoseangelroblesBelum ada peringkat
- Stem Cells: The Pursuit of Genomic Stability: Molecular SciencesDokumen20 halamanStem Cells: The Pursuit of Genomic Stability: Molecular SciencesjoseangelroblesBelum ada peringkat
- Yang 2007Dokumen13 halamanYang 2007joseangelroblesBelum ada peringkat
- To Manek 2015Dokumen13 halamanTo Manek 2015joseangelroblesBelum ada peringkat
- Sypro Ruby Staining Bio-Rad 4006173B-WMSDokumen10 halamanSypro Ruby Staining Bio-Rad 4006173B-WMSjoseangelroblesBelum ada peringkat
- Sauve 2003Dokumen8 halamanSauve 2003joseangelroblesBelum ada peringkat
- FunRichDocumentation 231214 V2.1.2Dokumen28 halamanFunRichDocumentation 231214 V2.1.2joseangelroblesBelum ada peringkat
- Plant Propagation Lab ActivityDokumen2 halamanPlant Propagation Lab ActivityClint Jhun TabugocBelum ada peringkat
- ABC Model of Flower Development - WikipediaDokumen9 halamanABC Model of Flower Development - WikipediaCarina JLBelum ada peringkat
- Science5 - q2 - Mod1 - The Parts of The Reproductive System and Their Functions - v4 PDFDokumen23 halamanScience5 - q2 - Mod1 - The Parts of The Reproductive System and Their Functions - v4 PDFLezel Laracas64% (11)
- The Aphrodisiac Activity of Ethanol Extract Bungkus Stem (Smilax Rotundifolia L.) On The Fertility of White Male Rats (Rattus Norvegicus)Dokumen10 halamanThe Aphrodisiac Activity of Ethanol Extract Bungkus Stem (Smilax Rotundifolia L.) On The Fertility of White Male Rats (Rattus Norvegicus)AyoeWoelandariBelum ada peringkat
- PDFDokumen4 halamanPDFmbakpithi0% (1)
- Repro Annotated First Aid 2019Dokumen51 halamanRepro Annotated First Aid 2019Heran TeferiBelum ada peringkat
- Paper 5 - CH - 7 Q. 1 HERIDITY AND ENVIRONMENTDokumen3 halamanPaper 5 - CH - 7 Q. 1 HERIDITY AND ENVIRONMENTnoorjhan dosaniBelum ada peringkat
- Paschoal & Zara (2020) - AABCDokumen19 halamanPaschoal & Zara (2020) - AABCLucas PaschoalBelum ada peringkat
- A Technology Enhanced Detailed Lesson Plan in Science 4 Designed by Panganiban, Baby Rica E. (BEED 2-2)Dokumen6 halamanA Technology Enhanced Detailed Lesson Plan in Science 4 Designed by Panganiban, Baby Rica E. (BEED 2-2)Monneth DalisayBelum ada peringkat
- 05 Pollen Spore ExaminationDokumen26 halaman05 Pollen Spore ExaminationRadu Ecaterina0% (1)
- Chapter 26Dokumen23 halamanChapter 26Konishko DeyBelum ada peringkat
- Stages of Lactation ExplainedDokumen35 halamanStages of Lactation ExplainedIgnacio FelicityBelum ada peringkat
- Yearly Plan For Science Form 3Dokumen19 halamanYearly Plan For Science Form 3Sue Suraya NazaBelum ada peringkat
- Ncma QADokumen9 halamanNcma QADANIELA DE GUZMANBelum ada peringkat
- Sex-Linked TraitsDokumen36 halamanSex-Linked TraitsRohini GagrooBelum ada peringkat
- Science: Quarter 2 - Module 1Dokumen20 halamanScience: Quarter 2 - Module 1Kate BatacBelum ada peringkat
- Compassion Without CompromiseDokumen30 halamanCompassion Without CompromiseBethany House PublishersBelum ada peringkat
- MaternalDokumen5 halamanMaternalStephanie Joy EscalaBelum ada peringkat
- Sci8 q4 Mod2 v4Dokumen31 halamanSci8 q4 Mod2 v4Jay-ar RiosBelum ada peringkat
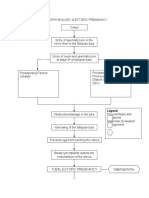
- PATHOPHYSIOLOGY of ECTOPIC PREGNANCYDokumen2 halamanPATHOPHYSIOLOGY of ECTOPIC PREGNANCYrye100% (1)
- Agrivita 2Dokumen10 halamanAgrivita 2Winda RachmadhaniBelum ada peringkat
- EU Reproduction Report Final ConsultationDokumen166 halamanEU Reproduction Report Final ConsultationMichael RhimesBelum ada peringkat
- Understanding Asexual vs Sexual ReproductionDokumen5 halamanUnderstanding Asexual vs Sexual ReproductionJah Jah Dalin Lpt100% (1)
- The POSEIDON Criteria and Its Measure of Success Through The Eyes of Clinicians and EmbryologistsDokumen8 halamanThe POSEIDON Criteria and Its Measure of Success Through The Eyes of Clinicians and EmbryologistsDimitrios PapadopoulosBelum ada peringkat
- Mode of ReproductionDokumen33 halamanMode of ReproductionMd. Rayhan Ali RejviBelum ada peringkat
- Test Questionnaires With Key Answer - Gabriel Aand BobisDokumen6 halamanTest Questionnaires With Key Answer - Gabriel Aand BobisLeah BobisBelum ada peringkat
- The REPRODUCTION-PROCESSDokumen30 halamanThe REPRODUCTION-PROCESSEJ GolimlimBelum ada peringkat
- Grade 9 Science (10F) : A Course For Independent StudyDokumen40 halamanGrade 9 Science (10F) : A Course For Independent StudyKing AlfonsoBelum ada peringkat
- Lesson 1 Stallion Management PowerpointDokumen25 halamanLesson 1 Stallion Management PowerpointelblazerBelum ada peringkat
- RotiferaDokumen12 halamanRotiferasuzayanarBelum ada peringkat