Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Joyway Std-3-19Dokumen40 halamanJoyway Std-3-19Vharry Watson33% (3)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Bobcat Classification and EvolutionDokumen3 halamanBobcat Classification and EvolutionagnaBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Ppt. Surgery Wild AnimalsDokumen165 halamanPpt. Surgery Wild AnimalsAsha RaniBelum ada peringkat
- Teddy Bear KeychainDokumen9 halamanTeddy Bear KeychainelaineribeBelum ada peringkat
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- British Expression DictionaryDokumen143 halamanBritish Expression DictionarySantoshBelum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Graded Reading: Animals in The City (Level 3) - Exercises: PreparationDokumen2 halamanGraded Reading: Animals in The City (Level 3) - Exercises: Preparationestefania aranda jimenezBelum ada peringkat
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Philippine LiteratureDokumen93 halamanPhilippine LiteratureEthel Marie Casido Burce100% (4)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Cesarean Section: Yusef C. Wibowo, S.ST.,M.HDokumen6 halamanCesarean Section: Yusef C. Wibowo, S.ST.,M.HberlyanaBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Brain Comparative Vertebrates Sem1Dokumen5 halamanThe Brain Comparative Vertebrates Sem1Rashmi SinghBelum ada peringkat
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- A Review Ot Esthetic Pontic Design OptionsDokumen12 halamanA Review Ot Esthetic Pontic Design OptionsAlejandro García Armenta100% (1)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Assesment C&G Prepare, Cook and Finish EggsDokumen6 halamanAssesment C&G Prepare, Cook and Finish Eggsعلي صالحBelum ada peringkat
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- E.V.O Search For Eden DetonadoDokumen51 halamanE.V.O Search For Eden DetonadoAlan KardecBelum ada peringkat
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Fate of The Embryonic Germ LayersDokumen3 halamanFate of The Embryonic Germ Layersmalav1989Belum ada peringkat
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Birds of Van ViharDokumen8 halamanBirds of Van ViharDurgesh Kumar SinghBelum ada peringkat
- Ã'Tia' Fiakdkdhl Úohd, H - FLD U 07: D.S. Senanayake College - Colombo 07Dokumen12 halamanÃ'Tia' Fiakdkdhl Úohd, H - FLD U 07: D.S. Senanayake College - Colombo 07joker boyBelum ada peringkat
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The 4x4 MatrixDokumen23 halamanThe 4x4 Matrixpfi_jenBelum ada peringkat
- Quarter 4 - CompleteDokumen115 halamanQuarter 4 - Completemaria luisa mendozaBelum ada peringkat
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Gagy Gygax's - Lejendary Adventure - Essentials Box Set - Moon SlavesDokumen13 halamanGagy Gygax's - Lejendary Adventure - Essentials Box Set - Moon Slavesestibandumas100% (1)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Describing FoodDokumen4 halamanDescribing FoodJohn SmithBelum ada peringkat
- The Scarecrow Queen (Excerpt)Dokumen36 halamanThe Scarecrow Queen (Excerpt)I Read YA50% (2)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Fairyland 1 LectiiDokumen7 halamanFairyland 1 LectiiLavinia SurmeiBelum ada peringkat
- Equus PDFDokumen8 halamanEquus PDFcfeauxBelum ada peringkat
- Physiology Final Exam - Glory 2017 PDFDokumen14 halamanPhysiology Final Exam - Glory 2017 PDFMohammad BarakatBelum ada peringkat
- GrammarDokumen12 halamanGrammarMuhamad Zaini MasryBelum ada peringkat
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Timesaver Grammar Activities Elem-PRESENTSIMPLE1Dokumen1 halamanTimesaver Grammar Activities Elem-PRESENTSIMPLE1Berta LlopisBelum ada peringkat
- CH 3 Read MeDokumen56 halamanCH 3 Read MeelzianhawkwindBelum ada peringkat
- Acute-Phase Protein - WikipediaDokumen4 halamanAcute-Phase Protein - WikipediaZACHARIAH MANKIRBelum ada peringkat
- Struktur Organisasi Puskesmas DoneDokumen19 halamanStruktur Organisasi Puskesmas Donepuskesmas kepulunganBelum ada peringkat
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
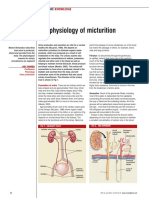
- 030722the Physiology of MicturitionDokumen3 halaman030722the Physiology of MicturitionAnnizah Paramitha100% (1)
- English TestDokumen13 halamanEnglish TestDJ MRBelum ada peringkat
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)