Anda mungkin juga menyukai
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Cultural Diffusion and The Kingdom of Kongo Essay OutlineDokumen3 halamanCultural Diffusion and The Kingdom of Kongo Essay OutlineAbenzer MamoBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5795)
- Arihant Kota Teachers - KVPY 11 Years Solved Papers 2019-2009 Stream SA For Class 11 Arihant (2020, Arihant Career Helper) - Libgen - LiDokumen257 halamanArihant Kota Teachers - KVPY 11 Years Solved Papers 2019-2009 Stream SA For Class 11 Arihant (2020, Arihant Career Helper) - Libgen - LiAtharv AggarwalBelum ada peringkat
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Review of Related LiteratureDokumen29 halamanReview of Related LiteratureTricsie VerdilloBelum ada peringkat
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Econometrics.: Home Assignment # 7Dokumen5 halamanEconometrics.: Home Assignment # 7Гая ЗимрутянBelum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Getting Things Done With ADHDDokumen74 halamanGetting Things Done With ADHDhfuqueneBelum ada peringkat
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- PCB Annual Report FormDokumen6 halamanPCB Annual Report FormNimshi TorreBelum ada peringkat
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Original PDF Introduction To Logic by Paul Herrick PDFDokumen41 halamanOriginal PDF Introduction To Logic by Paul Herrick PDFpearl.zinn592100% (35)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Rpa W3Dokumen15 halamanRpa W3MLCKB AMIRAH HAFIZAHBelum ada peringkat
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
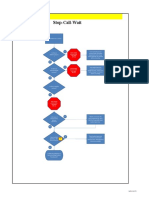
- Stop Call Wait BDokumen1 halamanStop Call Wait BTuan Anh100% (1)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Dimension EXL 200 Integrated Chemistry SystemDokumen3 halamanDimension EXL 200 Integrated Chemistry SystemNovan Pkbi100% (1)
- Theory of Thermal Energy He Atom Scattering From Ordered Submonolayers On Solid SurfacesDokumen136 halamanTheory of Thermal Energy He Atom Scattering From Ordered Submonolayers On Solid SurfacesAntonio ŠiberBelum ada peringkat
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Module 1 - Universities and National DevelopmentDokumen13 halamanModule 1 - Universities and National DevelopmentTOBI JASPER WANGBelum ada peringkat
- Production Logging Tools and InterpretationsDokumen34 halamanProduction Logging Tools and InterpretationsRizwan FaridBelum ada peringkat
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Liquid Crystal Colloidal Structures For IncreasedDokumen11 halamanLiquid Crystal Colloidal Structures For IncreasedCarlos Santos Bravo CcatamayoBelum ada peringkat
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Principles of Heating, Ventilation and Air Conditioning With Worked Examples PDFDokumen708 halamanPrinciples of Heating, Ventilation and Air Conditioning With Worked Examples PDFGBTBelum ada peringkat
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Chapter Three: Logic and LanguageDokumen16 halamanChapter Three: Logic and LanguageTagayBelum ada peringkat
- Disclosure To Promote The Right To InformationDokumen12 halamanDisclosure To Promote The Right To InformationShivashish ChaturvediBelum ada peringkat
- ConstellationsDokumen56 halamanConstellationsGelCess ParoanBelum ada peringkat
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Analysis of Single and Group Micropile Behavior PDFDokumen157 halamanAnalysis of Single and Group Micropile Behavior PDFLucas MartinsBelum ada peringkat
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Benefits and Side Effects of Baking Soda As A Deodorant.: Research ProposalDokumen9 halamanBenefits and Side Effects of Baking Soda As A Deodorant.: Research ProposalBea Lyn OrtegaBelum ada peringkat
- Some Guidelines and Guarantees For Common Random NumbersDokumen25 halamanSome Guidelines and Guarantees For Common Random NumbersAditya TBelum ada peringkat
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Testing Effects of Acid Rain On StatuesDokumen5 halamanTesting Effects of Acid Rain On StatuesDanielle LoneBelum ada peringkat
- Ra 5527Dokumen9 halamanRa 5527Sheena RojasBelum ada peringkat
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Pedu 202Dokumen44 halamanPedu 202Mejane MonterBelum ada peringkat
- Theater Arts Module 1Dokumen26 halamanTheater Arts Module 1yoonglespianoBelum ada peringkat
- Claim Applied Towards CapitationDokumen3 halamanClaim Applied Towards CapitationUavesh AnsariBelum ada peringkat
- Fed Question BankDokumen5 halamanFed Question BankAnonymous eWMnRr70qBelum ada peringkat
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- 英语个性化学程S1B,Unit 1-学生发放 PDFDokumen16 halaman英语个性化学程S1B,Unit 1-学生发放 PDFZi WangBelum ada peringkat
- Digital Unit PlanDokumen4 halamanDigital Unit Planapi-451554603Belum ada peringkat
- OPS Materials Specifications For JointsDokumen8 halamanOPS Materials Specifications For JointsKévin JacobBelum ada peringkat
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)