Anda mungkin juga menyukai
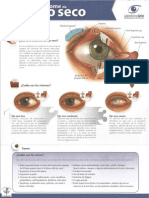
- Sindrome Ojo SecoDokumen2 halamanSindrome Ojo SecoDr. Javier Antonio Toledo LimónBelum ada peringkat
- Mercadotecnia Practica MedicaDokumen11 halamanMercadotecnia Practica MedicaDr. Javier Antonio Toledo LimónBelum ada peringkat
- Sindrome Tunel TarsianoDokumen5 halamanSindrome Tunel TarsianoDr. Javier Antonio Toledo LimónBelum ada peringkat
- Tratamiento Hipertension Arterial Sistemica Caso ClinicoDokumen15 halamanTratamiento Hipertension Arterial Sistemica Caso ClinicoDr. Javier Antonio Toledo Limón100% (1)
- Neuropatia Nervio MusculocutaneoDokumen7 halamanNeuropatia Nervio MusculocutaneoDr. Javier Antonio Toledo LimónBelum ada peringkat
- Infecciones Genitales No Complicadas Preguntas RespuestasDokumen9 halamanInfecciones Genitales No Complicadas Preguntas RespuestasDr. Javier Antonio Toledo LimónBelum ada peringkat
- CorticoterapiaDokumen25 halamanCorticoterapiaDr. Javier Antonio Toledo LimónBelum ada peringkat
- FibromialgiaDokumen7 halamanFibromialgiaDr. Javier Antonio Toledo Limón100% (1)
- Caso Clinico Diabetes Mellitus DescontroladoDokumen3 halamanCaso Clinico Diabetes Mellitus DescontroladoDr. Javier Antonio Toledo Limón0% (1)
- Comic Hiperplasia Prostatica BenignaDokumen15 halamanComic Hiperplasia Prostatica BenignaDr. Javier Antonio Toledo LimónBelum ada peringkat
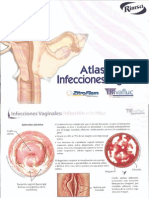
- Atlas Infecciones VaginalesDokumen10 halamanAtlas Infecciones VaginalesDr. Javier Antonio Toledo LimónBelum ada peringkat
- Caso Clinico Radiculopatia LumbarDokumen7 halamanCaso Clinico Radiculopatia LumbarDr. Javier Antonio Toledo LimónBelum ada peringkat
- Caso Clinico DM Hipertrigliceridemia Hiperlipidemia MixtaDokumen13 halamanCaso Clinico DM Hipertrigliceridemia Hiperlipidemia MixtaDr. Javier Antonio Toledo LimónBelum ada peringkat
- Caso Clinico Dermatitis SolarDokumen3 halamanCaso Clinico Dermatitis SolarDr. Javier Antonio Toledo LimónBelum ada peringkat
- Caso Clinico Dolor LumbarDokumen3 halamanCaso Clinico Dolor LumbarDr. Javier Antonio Toledo LimónBelum ada peringkat
- Caso Clinico Cistitis AgudaDokumen3 halamanCaso Clinico Cistitis AgudaDr. Javier Antonio Toledo LimónBelum ada peringkat
- El Bulbo RaquideoDokumen10 halamanEl Bulbo Raquideojulio roberto castilloBelum ada peringkat
- Tema 04. Ej - NerviosoDokumen5 halamanTema 04. Ej - NerviosoEstherOrtsSanchisBelum ada peringkat
- 10ma Clase Neuro - Sistematizacion SN - Dr. Pedro de La CruzDokumen47 halaman10ma Clase Neuro - Sistematizacion SN - Dr. Pedro de La CruzHinO - Arturo100% (1)
- Guia Tejido NeuromuscularDokumen7 halamanGuia Tejido NeuromuscularValentina RodriguezBelum ada peringkat
- IntroduccionsentidosDokumen31 halamanIntroduccionsentidosclicon22Belum ada peringkat
- AferenciasDokumen1 halamanAferenciasadrianBelum ada peringkat
- Organografía Microscópica HumanaDokumen144 halamanOrganografía Microscópica HumanaCristóbal Muñoz Pérez100% (2)
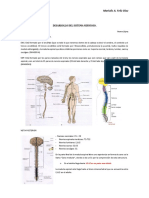
- Sistema Nervioso en El Hombre MCMDokumen9 halamanSistema Nervioso en El Hombre MCMProf. Víctor M. VitoriaBelum ada peringkat
- FisiologiaDokumen22 halamanFisiologiamajoBelum ada peringkat
- SX MedularesDokumen46 halamanSX MedularesFlores EleBelum ada peringkat
- Trabajo de Anatomia - Medula EspinalDokumen33 halamanTrabajo de Anatomia - Medula Espinalsilvia perez100% (1)
- Sistema Nervioso y EndocrinoDokumen39 halamanSistema Nervioso y EndocrinocarlabasbarBelum ada peringkat
- Sistema Nervioso 2Dokumen6 halamanSistema Nervioso 2CristianSantosBelum ada peringkat
- Funciones Motoras de La Medula EspinalDokumen31 halamanFunciones Motoras de La Medula EspinalLuis Felipe CarrilloBelum ada peringkat
- Todos Los FHFDokumen7 halamanTodos Los FHFcamilisangelicaBelum ada peringkat
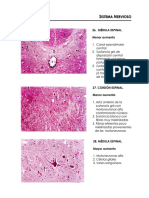
- La Médula EspinalDokumen11 halamanLa Médula EspinalCristofer Abanto García100% (2)
- Tarea No.3 Sistema NerviosoDokumen6 halamanTarea No.3 Sistema NerviosoDiego AlvaradoBelum ada peringkat
- Clases en Word Teoria Primer ParcialDokumen70 halamanClases en Word Teoria Primer ParcialViangely SantanaBelum ada peringkat
- Tema 8 Sistema NerviosoDokumen8 halamanTema 8 Sistema NerviosoMONICA SALMERON MONTEROBelum ada peringkat
- Informe, MédulaDokumen4 halamanInforme, MédulaJARBelum ada peringkat
- Sistemas - Sensitivos - Generales 2Dokumen11 halamanSistemas - Sensitivos - Generales 2Jimena Reyes0% (1)
- SNCDokumen37 halamanSNCMiguel Ángel TórrezBelum ada peringkat
- Analgesia Por AcupunturaDokumen36 halamanAnalgesia Por AcupunturaJonathan100% (1)
- GUIAS-MORFOLOGÍA-I-HISTOLOGÍA-AZ-SIGNIFICATIVO-2-2019-sin TegDokumen36 halamanGUIAS-MORFOLOGÍA-I-HISTOLOGÍA-AZ-SIGNIFICATIVO-2-2019-sin TegpaolaBelum ada peringkat
- Lesión Medular Trabajo GrupalDokumen1 halamanLesión Medular Trabajo GrupalEralda HushBelum ada peringkat
- IntroducciónDokumen49 halamanIntroducciónAlejandro MCBelum ada peringkat
- Funciones Motoras de La Medula Espinal Reflejos Medulares DR MerelloDokumen83 halamanFunciones Motoras de La Medula Espinal Reflejos Medulares DR Merellojosemerello100% (1)
- Semana 10Dokumen15 halamanSemana 10CAMILA VALERIABelum ada peringkat
- Trauma RaquimedularDokumen42 halamanTrauma RaquimedularMadeleine Louvre100% (1)
- Anatomia y Fisiologia ME 2Dokumen73 halamanAnatomia y Fisiologia ME 2javier fernandezBelum ada peringkat