Anda mungkin juga menyukai
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Drug Study - Anesthestics....Dokumen43 halamanDrug Study - Anesthestics....Edward Baes67% (3)
- 1 MSN UNIT 1introductionDokumen57 halaman1 MSN UNIT 1introductionVikash PrajapatiBelum ada peringkat
- Reviewer (Gen. Biology 2)Dokumen5 halamanReviewer (Gen. Biology 2)ianah aloriaBelum ada peringkat
- First Announcement PIN IKABDI Medan 2024Dokumen11 halamanFirst Announcement PIN IKABDI Medan 2024pinikabdimedan2024100% (1)
- Diabetes Advances in Diagnosis and TreatmentDokumen11 halamanDiabetes Advances in Diagnosis and TreatmentAbi GarciaBelum ada peringkat
- Hiking Concept and BenefitsDokumen30 halamanHiking Concept and BenefitsSteven TaperoBelum ada peringkat
- Types, Symptoms, and Treatment of Kidney StonesDokumen6 halamanTypes, Symptoms, and Treatment of Kidney StonesKatte VillenasBelum ada peringkat
- Unit I Introduction To HealthDokumen11 halamanUnit I Introduction To Healthishir babu sharmaBelum ada peringkat
- Craniofacial Surgery Dx and Tx TechniquesDokumen103 halamanCraniofacial Surgery Dx and Tx TechniquesSiddharth DhanarajBelum ada peringkat
- Mastering the Medical Long Case: A Guide for Physician TraineesDokumen20 halamanMastering the Medical Long Case: A Guide for Physician TraineesHasak3Belum ada peringkat
- First AidDokumen4 halamanFirst Aidrebz100% (1)
- ReportDokumen21 halamanReportRenz CruzBelum ada peringkat
- Reading Tips TOEFLDokumen34 halamanReading Tips TOEFLFitri PuspasariBelum ada peringkat
- HypertensionDokumen19 halamanHypertensionmr_coolz282344Belum ada peringkat
- DADHDokumen69 halamanDADHdrpnnreddyBelum ada peringkat
- Rome III Diagnostic Criteria FGIDsDokumen14 halamanRome III Diagnostic Criteria FGIDsPutu Reza Sandhya PratamaBelum ada peringkat
- Vancomycin - 3rd Generation GlycopeptideDokumen3 halamanVancomycin - 3rd Generation GlycopeptideJenny Agustin FabrosBelum ada peringkat
- Family Medical TreeDokumen2 halamanFamily Medical TreeDevi MayasariBelum ada peringkat
- Tuberculosis / TBDokumen11 halamanTuberculosis / TBSherree HayesBelum ada peringkat
- MCQ 1Dokumen4 halamanMCQ 1Maxine foyeBelum ada peringkat
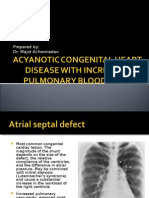
- Acyanotic Congenital Heart Disease With Increased Pulmonary Blood FlowDokumen9 halamanAcyanotic Congenital Heart Disease With Increased Pulmonary Blood Flowdrhomiedan100% (1)
- Radiologia en EndodonciaDokumen12 halamanRadiologia en EndodonciaBelez Jum JumBelum ada peringkat
- Manzolli Etal 2015Dokumen8 halamanManzolli Etal 2015gustavormbarcelosBelum ada peringkat
- Medical Nursing 1 UpdatedDokumen148 halamanMedical Nursing 1 UpdatedcomfortaporoBelum ada peringkat
- What is GMO? The Complete GuideDokumen7 halamanWhat is GMO? The Complete GuidewerkaserkaBelum ada peringkat
- Myasthenia GravisDokumen11 halamanMyasthenia Gravishironbose0% (1)
- 0 - Hydrogen Water Generator Mini (Eng) - India Version - Read OnlyDokumen24 halaman0 - Hydrogen Water Generator Mini (Eng) - India Version - Read OnlyPrakash Aradhya R100% (2)
- F QuesPaper INBO2012Dokumen39 halamanF QuesPaper INBO2012martynapet100% (1)
- Bishop Score - StatPearls - NCBI BookshelfDokumen3 halamanBishop Score - StatPearls - NCBI BookshelfDewa ayu NarheswariBelum ada peringkat
- The Tamil Nadu Dr. M.G.R. Medical University - MSC Nursing Second YrDokumen10 halamanThe Tamil Nadu Dr. M.G.R. Medical University - MSC Nursing Second YrNoufal100% (1)