Anda mungkin juga menyukai
- Apuntes de Anatomía de La MaderaDokumen13 halamanApuntes de Anatomía de La MaderaNathalie Le NoirBelum ada peringkat
- Anatomia y Fisiologia VegetalDokumen15 halamanAnatomia y Fisiologia VegetalDaiana OlimaBelum ada peringkat
- El Tejido Epidérmico Vegetal Es El Protector Vivo Que Recubre La Superficie de Toda La Planta Cuando Ésta Posee Estructura PrimariaDokumen9 halamanEl Tejido Epidérmico Vegetal Es El Protector Vivo Que Recubre La Superficie de Toda La Planta Cuando Ésta Posee Estructura PrimariaValeriaDlcoBelum ada peringkat
- Tejidos VegetalesDokumen8 halamanTejidos VegetalesMaria Alejandra Rodriguez EspinozaBelum ada peringkat
- Resumen de Tejidos VegetalesDokumen3 halamanResumen de Tejidos VegetalesCarolBelum ada peringkat
- Practica de AnatomíaDokumen26 halamanPractica de AnatomíaMarcos OvalleBelum ada peringkat
- Tejidos VegetalesDokumen8 halamanTejidos Vegetalesbetincarlos3010Belum ada peringkat
- Anatomia de Los Tejidos VegetalesDokumen9 halamanAnatomia de Los Tejidos VegetaleslollihopBelum ada peringkat
- BOTÁNICADokumen46 halamanBOTÁNICAheccf000008579Belum ada peringkat
- Tejidos VegetalesDokumen5 halamanTejidos Vegetalesiris chirinosBelum ada peringkat
- Célula VegetalDokumen2 halamanCélula VegetalNataly MoralesBelum ada peringkat
- Tejidos VegetalesDokumen9 halamanTejidos VegetalesEner Vilchez CuevasBelum ada peringkat
- Transformacion CelularDokumen24 halamanTransformacion CelularbiotechdogoBelum ada peringkat
- Cuáles Son Los Tipos de Tejidos VegetalesDokumen15 halamanCuáles Son Los Tipos de Tejidos VegetalespaulaingenieriacivilBelum ada peringkat
- Botánica - Tema 06Dokumen23 halamanBotánica - Tema 06Perez BalcarceBelum ada peringkat
- Tejidos VegetalesDokumen4 halamanTejidos VegetalesMary PeñalverBelum ada peringkat
- Resumen de Tejidos VegetalesDokumen10 halamanResumen de Tejidos VegetalesRoland M ReyesBelum ada peringkat
- Tejido Sostén-ConductoresDokumen6 halamanTejido Sostén-ConductoresAbigailBelum ada peringkat
- Tejidos EmbrionariosDokumen4 halamanTejidos EmbrionariosAle Al Cubo100% (1)
- Tejidos de SostenDokumen5 halamanTejidos de SostenCarlos Alexander L. HernandezBelum ada peringkat
- Tejidos Vegetales 6 y 7Dokumen6 halamanTejidos Vegetales 6 y 7lorenzo quezadaBelum ada peringkat
- Laboratorio Tejidos Vegetales Jhefrey AngaritaDokumen8 halamanLaboratorio Tejidos Vegetales Jhefrey Angaritajere angaritaBelum ada peringkat
- Definición de BiologíaDokumen4 halamanDefinición de BiologíaTC DacneBelum ada peringkat
- Tejidos Epidemico, Parenquimatico, Conductor, Suberoso, MeristematicoDokumen6 halamanTejidos Epidemico, Parenquimatico, Conductor, Suberoso, MeristematicoCecy GuerraBelum ada peringkat
- Tejido VegetalDokumen7 halamanTejido VegetalCarlosrojasbarriosBelum ada peringkat
- Tejidos VegetalesDokumen16 halamanTejidos Vegetalesphysicx1725Belum ada peringkat
- Guia 4 Tejidos VegetalesDokumen9 halamanGuia 4 Tejidos VegetalesEsperanza YunithBelum ada peringkat
- Enfoque de La Célula Vegetal..Dokumen12 halamanEnfoque de La Célula Vegetal..José DaríoBelum ada peringkat
- Celula y Tejido VegetalDokumen46 halamanCelula y Tejido VegetalWadir Flores tisnadoBelum ada peringkat
- CuestionarioDokumen8 halamanCuestionarioErick AcostaBelum ada peringkat
- Tema 15 - Tejidos Vegetales IDokumen9 halamanTema 15 - Tejidos Vegetales ISandra Díez AlkaldeBelum ada peringkat
- Tejido Vegetal HistologíaDokumen11 halamanTejido Vegetal HistologíaasenfhnawfBelum ada peringkat
- Los Tegidos VegetalesDokumen15 halamanLos Tegidos Vegetalesrosalinrosalesc100% (2)
- Tej VegDokumen4 halamanTej VegbiologorogerBelum ada peringkat
- T.12: Tejido Fundamental Parénquima. Tejido de Sostén Esteroma, Colénquima Y EsclerénquimaDokumen3 halamanT.12: Tejido Fundamental Parénquima. Tejido de Sostén Esteroma, Colénquima Y Esclerénquimavoldur12Belum ada peringkat
- Organización Celular de Las PlantasDokumen57 halamanOrganización Celular de Las PlantasRaúl Dylan CapchaBelum ada peringkat
- Parcial II - BotánicaDokumen25 halamanParcial II - BotánicaPerez BalcarceBelum ada peringkat
- TALLER 1 BiologiaDokumen6 halamanTALLER 1 BiologiaJESUS DANIEL VILLAMIZAR MONTOYABelum ada peringkat
- Biologia 025 Tejidos VegetalesDokumen10 halamanBiologia 025 Tejidos VegetalesMoroni45Belum ada peringkat
- HISTOLOGIADokumen2 halamanHISTOLOGIACristhian Fernando Crespo MuñozBelum ada peringkat
- Tejidos de Las PlantasDokumen20 halamanTejidos de Las PlantasAndrea Acevedo PeñalozaBelum ada peringkat
- 05.1 Tejido ProtectorDokumen91 halaman05.1 Tejido ProtectorYeferson RossellóBelum ada peringkat
- Informe de Fisologia VegetalDokumen14 halamanInforme de Fisologia VegetalBrat AcuacheBelum ada peringkat
- Anatomia y Fisiologia Vegetal (1) .... Tjidos..SemillasDokumen38 halamanAnatomia y Fisiologia Vegetal (1) .... Tjidos..Semillasviktor26Belum ada peringkat
- Práctica 2 Célula y Tejidos VegetalesDokumen14 halamanPráctica 2 Célula y Tejidos VegetalesPerezortiz 18 GerardoBelum ada peringkat
- Proteccion 2Dokumen17 halamanProteccion 2Camila EsRaBelum ada peringkat
- Anatomía y Fisiología Vegetal IDokumen5 halamanAnatomía y Fisiología Vegetal IGriselita RamirezBelum ada peringkat
- Biologia DeberDokumen4 halamanBiologia DeberAracely HerreraBelum ada peringkat
- Clasificación de Los Teejidos VegetalDokumen11 halamanClasificación de Los Teejidos VegetalAlex Pimentel Donayre100% (1)
- Histología VegetalDokumen9 halamanHistología VegetalJuan ZuñigaBelum ada peringkat
- La Celula VegetalDokumen9 halamanLa Celula VegetalRoberto AndersonBelum ada peringkat
- Practica 3Dokumen4 halamanPractica 3Emerson VargasBelum ada peringkat
- Tejidos de SoporteDokumen9 halamanTejidos de SoporteJuan Ariel Aspi ApazaBelum ada peringkat
- Guia 1 MeristemasDokumen3 halamanGuia 1 MeristemasCatalina CfBelum ada peringkat
- Estructura y Composicion Del Tejido Vegetal PresentacionDokumen40 halamanEstructura y Composicion Del Tejido Vegetal Presentacionalexander naspiranBelum ada peringkat
- Unidad 07 EpidermisDokumen5 halamanUnidad 07 EpidermisLautaro TournBelum ada peringkat
- GlosarioDokumen13 halamanGlosarioJocelyne Calderon PadillaBelum ada peringkat
- BC-Seminarios 1-2020-2021Dokumen6 halamanBC-Seminarios 1-2020-2021SanBor 02Belum ada peringkat
- Acuerdo 434-12 Resolucion de ConflictosDokumen5 halamanAcuerdo 434-12 Resolucion de ConflictosCesar Carlos Cervantes50% (2)
- Solicitud de Acumulacion de DecimosDokumen1 halamanSolicitud de Acumulacion de DecimosLianita MorenaBelum ada peringkat
- El AlcoholismoDokumen25 halamanEl AlcoholismoAngelica Romo TijerinaBelum ada peringkat
- Exposicion Educacion Ambiental Sindicato de Choferes QuezadaDokumen24 halamanExposicion Educacion Ambiental Sindicato de Choferes QuezadaLianita Morena100% (2)
- Formicidae de MexicoDokumen36 halamanFormicidae de MexicoFernandaBelum ada peringkat
- Mi Libro de Tareas Preescolar. Star Creando. Star LeyvaDokumen104 halamanMi Libro de Tareas Preescolar. Star Creando. Star LeyvaNallely Guzwan Zanchez100% (1)
- Mentefacto GrupalDokumen3 halamanMentefacto Grupalariel_camilo2000Belum ada peringkat
- Cuadrantes Del AbdomenDokumen36 halamanCuadrantes Del AbdomenAlejandra EstupiñanBelum ada peringkat
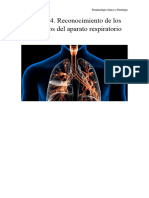
- Sistema Respiratorio CamilaDokumen7 halamanSistema Respiratorio CamilaKarina Itzamary Cadenas100% (1)
- Mapa Conceptual Del Sistema CirculatorioDokumen1 halamanMapa Conceptual Del Sistema CirculatorioMyriam Gladys Pruna OrozcoBelum ada peringkat
- Seminario 1 Tejido EpitelialDokumen5 halamanSeminario 1 Tejido EpitelialLaura MatosBelum ada peringkat
- 8 - Sangre y HematogenesisDokumen45 halaman8 - Sangre y HematogenesisJosue GarciaBelum ada peringkat
- Las Lampreas.Dokumen1 halamanLas Lampreas.Rubi Enrriquez DecenaBelum ada peringkat
- Selva Del CongoDokumen32 halamanSelva Del Congoashley peñaBelum ada peringkat
- Partes Del Aparato Digestivo y Sus FuncionesDokumen10 halamanPartes Del Aparato Digestivo y Sus FuncionesprobiiBelum ada peringkat
- Adolescencia InicialDokumen2 halamanAdolescencia InicialMathias CoelloBelum ada peringkat
- Locomocion Humana FASE 2 UDokumen20 halamanLocomocion Humana FASE 2 UGelnys Alonso Vasquez CuevasBelum ada peringkat
- Prueba de CienciasDokumen3 halamanPrueba de Ciencias19081512alanalluBelum ada peringkat
- Evaluación de Ciencias Naturales 5° Sistema Respiratorio y CirculatorioDokumen2 halamanEvaluación de Ciencias Naturales 5° Sistema Respiratorio y CirculatorioChinita Soledad Muñoz Garrido100% (1)
- Dermis. Eq7. g1351Dokumen16 halamanDermis. Eq7. g1351Marlene BlackBelum ada peringkat
- HELMINTOSDokumen8 halamanHELMINTOSErik J. Roshan FloresBelum ada peringkat
- Tema 3 EmbriologiaDokumen8 halamanTema 3 EmbriologiaJulio SalazarBelum ada peringkat
- Sistema EndocrinoDokumen18 halamanSistema EndocrinoephifanniaBelum ada peringkat
- Miotomas y DermatomasDokumen9 halamanMiotomas y DermatomasYazmin Rosales MimbelaBelum ada peringkat
- Sistemas Del CuerpoDokumen5 halamanSistemas Del CuerpoMaria PazBelum ada peringkat
- MIOLOGIADokumen15 halamanMIOLOGIAAnonymous Ta8Yls2Belum ada peringkat
- Los Peces. Introducción A Su Anatomía.Dokumen164 halamanLos Peces. Introducción A Su Anatomía.NoeBelum ada peringkat
- Plan LectorDokumen2 halamanPlan LectorAder Cachi QuispeBelum ada peringkat
- Cavidad Abdominal. EXPO ANATODokumen11 halamanCavidad Abdominal. EXPO ANATOSebastian RangelBelum ada peringkat
- Ejercicios Unidad4 TerminologiaDokumen7 halamanEjercicios Unidad4 TerminologiaDaniel MudarraBelum ada peringkat
- Taller 1 Biologia 8º: Sistema NerviosoDokumen6 halamanTaller 1 Biologia 8º: Sistema NerviosoCarlos BBelum ada peringkat
- Tejido OseoDokumen1 halamanTejido OseoMarlene SaucedoBelum ada peringkat
- Evaluación de La Vía Aérea Superior en OrtodonciaDokumen1 halamanEvaluación de La Vía Aérea Superior en OrtodonciaJhon CastroBelum ada peringkat
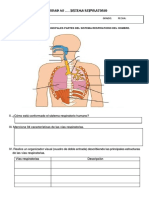
- Ficha de Trabajo Sistema RespiratorioDokumen5 halamanFicha de Trabajo Sistema Respiratoriodoris flores colana100% (2)