Anda mungkin juga menyukai
- Test Bank For Lehninger Principles of Biochemistry Seventh EditionDokumen24 halamanTest Bank For Lehninger Principles of Biochemistry Seventh Editioncosimalocu68xb1Belum ada peringkat
- Cell SignallingDokumen12 halamanCell SignallingRibka Kristania HadhiwaluyoBelum ada peringkat
- Management of Aging SkinDokumen19 halamanManagement of Aging SkinmonitamiftahBelum ada peringkat
- MCQs Series For Life SciencesDokumen579 halamanMCQs Series For Life Sciencesshwetha100% (2)
- Physical ChemistryDokumen43 halamanPhysical Chemistrydrami94Belum ada peringkat
- Respiratory Chain & Oxidative PhosphorylationDokumen57 halamanRespiratory Chain & Oxidative PhosphorylationHanifa AffianiBelum ada peringkat
- Unit 5 Developmental Biology CSIR UGC NET Life Sciences PDFDokumen5 halamanUnit 5 Developmental Biology CSIR UGC NET Life Sciences PDFjaspreet kaurBelum ada peringkat
- Chapter 8 - Nucleotides and Nucleic Acids - Notes For StudentsDokumen4 halamanChapter 8 - Nucleotides and Nucleic Acids - Notes For StudentsDjZeus1100% (1)
- 01.coordination Chemistry Class Notes Part I-1 PDFDokumen86 halaman01.coordination Chemistry Class Notes Part I-1 PDFShadrack Peter100% (1)
- Cell Biology and Membrane BiochemistryDokumen106 halamanCell Biology and Membrane BiochemistryVijay Bhasker TekulapallyBelum ada peringkat
- RAO Molecular BiologyDokumen113 halamanRAO Molecular BiologyH.J.PrabhuBelum ada peringkat
- Biomolecules: Amino Acids, Peptides, and ProteinsDokumen55 halamanBiomolecules: Amino Acids, Peptides, and ProteinsBoooooooseBelum ada peringkat
- Biochemistry TymockzcoDokumen315 halamanBiochemistry Tymockzcoswaala4realBelum ada peringkat
- Chapter 1 Biochemistry and The Organization of CellsDokumen14 halamanChapter 1 Biochemistry and The Organization of Cellskeziazah100% (1)
- Cell SignalingDokumen6 halamanCell SignalingEmBelum ada peringkat
- Biochemistry Student BookDokumen69 halamanBiochemistry Student BookWahed WatanyBelum ada peringkat
- Multiple Choice Questions in Molecular BiologyDokumen72 halamanMultiple Choice Questions in Molecular Biologydanishuddin_danish50% (2)
- Encyclopedia of Inorganic Chemistry PDFDokumen191 halamanEncyclopedia of Inorganic Chemistry PDFDan Bo100% (2)
- Lec 7 Biogeochemical CyclesDokumen12 halamanLec 7 Biogeochemical CyclesKhaled Hasan Khan100% (1)
- Chapter Three Amino Acids and Peptides: Paul D. Adams - University of ArkansasDokumen27 halamanChapter Three Amino Acids and Peptides: Paul D. Adams - University of ArkansasSheila GarciaBelum ada peringkat
- Lehninger LCA Ch03Dokumen87 halamanLehninger LCA Ch03Gavin100% (1)
- Lehninger LCA Ch01Dokumen81 halamanLehninger LCA Ch01Gavin100% (1)
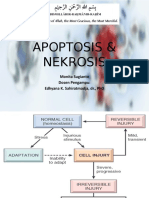
- Apoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDDokumen34 halamanApoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDmonitamiftahBelum ada peringkat
- LaserDokumen104 halamanLasermonitamiftah100% (1)
- Nucleic Acids 2015Dokumen72 halamanNucleic Acids 2015meh100% (1)
- Lehninger LCA Ch02Dokumen44 halamanLehninger LCA Ch02GavinBelum ada peringkat
- BiochemistryDokumen47 halamanBiochemistryFaizaAnum100% (1)
- Lehninger PPT ch04Dokumen73 halamanLehninger PPT ch04MahdeeHaqueSyed100% (1)
- In-Depth Steps Towards Nucleic Acid and Protein SynthesisDokumen21 halamanIn-Depth Steps Towards Nucleic Acid and Protein SynthesisGbenga AjaniBelum ada peringkat
- CH 3 - ProblemsDokumen6 halamanCH 3 - ProblemsKhris Griffis100% (5)
- Practical BiochemistryDokumen108 halamanPractical BiochemistryCindy Nona100% (1)
- FST Undergraduate Handbook 2020-2021Dokumen392 halamanFST Undergraduate Handbook 2020-2021Robin WilliamsBelum ada peringkat
- Environmental BiotechnologyDokumen130 halamanEnvironmental Biotechnologyshivnetrasrk100% (3)
- Lehninger LCA Ch06Dokumen113 halamanLehninger LCA Ch06GavinBelum ada peringkat
- Inflammation in AgingDokumen14 halamanInflammation in AgingmonitamiftahBelum ada peringkat
- SunscreenDokumen8 halamanSunscreenmonitamiftahBelum ada peringkat
- Botney Notes of 1,2,4 ChapterDokumen69 halamanBotney Notes of 1,2,4 ChapterJavEria Urooj DanWarBelum ada peringkat
- Biochemistry Test Bank ch1 (Fundamental of Biochemistry)Dokumen2 halamanBiochemistry Test Bank ch1 (Fundamental of Biochemistry)Ibrahim BarhamBelum ada peringkat
- Components in Fats and OilsDokumen18 halamanComponents in Fats and Oilsruman ahmad100% (1)
- Content Lehninger Biochemistry BookDokumen2 halamanContent Lehninger Biochemistry BookPingkan Liana MaharaniBelum ada peringkat
- Indian Constituition 1 WordsDokumen9 halamanIndian Constituition 1 WordsChinnasamy100% (1)
- LifeSciencesPart-1 FifthEditionDokumen176 halamanLifeSciencesPart-1 FifthEditionMuhammad Usman AkramBelum ada peringkat
- BiochemistryDokumen113 halamanBiochemistryMohammed Faizuddin siddiqui100% (1)
- Sterols 190319Dokumen29 halamanSterols 190319Sathish Sizzy0% (1)
- 5 KOF Acid Bases PDFDokumen27 halaman5 KOF Acid Bases PDFIstiBelum ada peringkat
- Environmental BiotechnologyDokumen13 halamanEnvironmental BiotechnologydgkulkarniBelum ada peringkat
- Biomolecules Reading Sample Part 1-Edited For NotesDokumen2 halamanBiomolecules Reading Sample Part 1-Edited For Notesapi-327757745Belum ada peringkat
- Experiment No. 12 Nucleic AcidsDokumen4 halamanExperiment No. 12 Nucleic AcidsJulia IgnacioBelum ada peringkat
- 202 Biochemistry PracticalsDokumen2 halaman202 Biochemistry PracticalsStephen MooreBelum ada peringkat
- Multiple Choice Questions-Cancers: AdvertisementDokumen5 halamanMultiple Choice Questions-Cancers: AdvertisementSp PpvBelum ada peringkat
- Molecular Biology ExamDokumen8 halamanMolecular Biology Exam畏Belum ada peringkat
- Pharm D SyllabusDokumen120 halamanPharm D SyllabusDR Salih Ibrahim60% (5)
- Fischer Projection FormulasDokumen2 halamanFischer Projection Formulasepah_05Belum ada peringkat
- Techniques of Molecular Biology, The Polymerase Chain Reaction (PCR)Dokumen22 halamanTechniques of Molecular Biology, The Polymerase Chain Reaction (PCR)Dr. Atif Hassan Khirelsied100% (1)
- Molecular BiotechnologyDokumen40 halamanMolecular BiotechnologyBhaskar GangulyBelum ada peringkat
- Med Chem Exam 2Dokumen24 halamanMed Chem Exam 2cBelum ada peringkat
- 3 Genetics: 3.1 GenesDokumen22 halaman3 Genetics: 3.1 Genesapi-285490927100% (1)
- Molecular and Cell Biology QuizzesDokumen60 halamanMolecular and Cell Biology QuizzesTyler Harvey100% (8)
- 5BBB0223 Mitochondrial Function & Dysfunction: Timothy Pullen With Thanks To: DR Clare Thornton, RVCDokumen63 halaman5BBB0223 Mitochondrial Function & Dysfunction: Timothy Pullen With Thanks To: DR Clare Thornton, RVCW BBelum ada peringkat
- Oxidative Phosphorylation and Electron Transport Chain II Respiratory Complexes Structure and Organization Chemiosmotic TheoryDokumen20 halamanOxidative Phosphorylation and Electron Transport Chain II Respiratory Complexes Structure and Organization Chemiosmotic TheoryIffatnazBelum ada peringkat
- Cellular RespirationDokumen55 halamanCellular RespirationFuad sabsebBelum ada peringkat
- Electron Transport SystemDokumen58 halamanElectron Transport SystemSantosh KumarBelum ada peringkat
- Tema 6quim Al17-18Dokumen39 halamanTema 6quim Al17-18noelia100997Belum ada peringkat
- Electrontransportchain 151027073626 Lva1 App6892Dokumen29 halamanElectrontransportchain 151027073626 Lva1 App6892GobindaSahuBelum ada peringkat
- Gayatree PaniDokumen23 halamanGayatree PaniGobindaSahuBelum ada peringkat
- Oral Collagen Supplementation For Skin Aging: A Fad or The Future?Dokumen3 halamanOral Collagen Supplementation For Skin Aging: A Fad or The Future?monitamiftahBelum ada peringkat
- Lavees Cosmedics: Cara PakaiDokumen2 halamanLavees Cosmedics: Cara PakaimonitamiftahBelum ada peringkat
- Asam Traneksamat Untuk KulitDokumen2 halamanAsam Traneksamat Untuk KulitmonitamiftahBelum ada peringkat
- Seminar: Heidi D NelsonDokumen11 halamanSeminar: Heidi D NelsonmonitamiftahBelum ada peringkat
- Guidelines Acne Vulgaris PDFDokumen14 halamanGuidelines Acne Vulgaris PDFmonitamiftahBelum ada peringkat
- Apoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDDokumen34 halamanApoptosis & Nekrosis: Monita Sugianto Dosen Pengampu: Edhyana K. Sahiratmadja, DR., PHDmonitamiftahBelum ada peringkat
- Reviews: Immunity To Fungal InfectionsDokumen13 halamanReviews: Immunity To Fungal InfectionsmonitamiftahBelum ada peringkat
- Efek Immunologi Radiasi Sinar UV MatahariDokumen16 halamanEfek Immunologi Radiasi Sinar UV MataharimonitamiftahBelum ada peringkat
- Efek Immunologi Radiasi Sinar UV MatahariDokumen16 halamanEfek Immunologi Radiasi Sinar UV MataharimonitamiftahBelum ada peringkat