Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Occupational Hazards in Health Care ProfessionalsDokumen33 halamanOccupational Hazards in Health Care Professionalsdocshirin100% (1)
- Minimalist Shoes Running Intervention Can Alter The Plantar Loadingdistribution and Deformation of Hallux Valgus A Pilot StudyDokumen7 halamanMinimalist Shoes Running Intervention Can Alter The Plantar Loadingdistribution and Deformation of Hallux Valgus A Pilot StudyGourgen GevorgyanBelum ada peringkat
- Degnala DiseaseDokumen12 halamanDegnala DiseaseSantosh Bhandari100% (1)
- (Godefroy Christian) Super Health PDFDokumen231 halaman(Godefroy Christian) Super Health PDFAlexandre Dantas100% (1)
- KetamineDokumen4 halamanKetamineapi-235915759Belum ada peringkat
- 95 Formulation and Evaluation of Diclofenac Sodium Gel by Using Natural PolymerDokumen3 halaman95 Formulation and Evaluation of Diclofenac Sodium Gel by Using Natural PolymerJulian Kayne100% (1)
- Limfoma Kutis Pada Pasien Yang Semula Di DiagnosisDokumen34 halamanLimfoma Kutis Pada Pasien Yang Semula Di DiagnosisChris MulyoBelum ada peringkat
- Tushar FinalDokumen29 halamanTushar FinalRaj Prixit RathoreBelum ada peringkat
- Soccer Programme U19 Pre in SeasonDokumen38 halamanSoccer Programme U19 Pre in SeasonJeffrey Peeko100% (1)
- Client Choice Plus Key FactsDokumen4 halamanClient Choice Plus Key Factsjudge101Belum ada peringkat
- Budwig Cancer GuideDokumen101 halamanBudwig Cancer Guidedora.oreiro9920Belum ada peringkat
- Concept Map 3 MM (Abdominal Pain)Dokumen2 halamanConcept Map 3 MM (Abdominal Pain)Matt McKinleyBelum ada peringkat
- Treatment of Oropharyngeal Dysphagia CarolMcKee (1) DPNS StudyDokumen23 halamanTreatment of Oropharyngeal Dysphagia CarolMcKee (1) DPNS Studyconstantineelia100% (1)
- Imperforate Anus PDFDokumen2 halamanImperforate Anus PDFGlyneth FranciaBelum ada peringkat
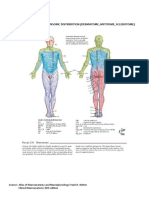
- Dermatome Myotome SclerotomeDokumen4 halamanDermatome Myotome SclerotomeAdhya TiaraBelum ada peringkat
- Damolo, Jules Aldrich C. October 30, 2021 Bsit 1 Year Block - A Instructor: Jerson CamayDokumen3 halamanDamolo, Jules Aldrich C. October 30, 2021 Bsit 1 Year Block - A Instructor: Jerson CamayAldrich Dos DamoloBelum ada peringkat
- Introduction To SpineDokumen29 halamanIntroduction To Spinebmahmood1Belum ada peringkat
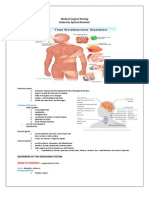
- Endocrine SystemDokumen8 halamanEndocrine Systemtheglobalnursing80% (5)
- Nasya Treatment: DefinitionDokumen4 halamanNasya Treatment: DefinitionkaukabBelum ada peringkat
- Long-Term Prediction of Functional Outcome After Stroke Using Single Items of The Barthel Index at Discharge From Rehabilitation CentreDokumen7 halamanLong-Term Prediction of Functional Outcome After Stroke Using Single Items of The Barthel Index at Discharge From Rehabilitation CentretitinBelum ada peringkat
- Introduction To Anatomy and Homeostasis Within The BodyDokumen24 halamanIntroduction To Anatomy and Homeostasis Within The BodyTrinity SherBelum ada peringkat
- Bibliografia 2222Dokumen28 halamanBibliografia 2222francivan111Belum ada peringkat
- Brodil LevoDokumen2 halamanBrodil LevoAziza KhairunBelum ada peringkat
- Guiding Principles For Best Practices in Geriatric PTDokumen16 halamanGuiding Principles For Best Practices in Geriatric PTPritesh KujurBelum ada peringkat
- Teratogenic ItyDokumen8 halamanTeratogenic ItyValentin IanaBelum ada peringkat
- Family Health Problem Asthma As Health DeficitDokumen3 halamanFamily Health Problem Asthma As Health Deficitiloveanne75% (4)
- Patient Feedback Form Fillable Version 2Dokumen2 halamanPatient Feedback Form Fillable Version 2GauravChopraBelum ada peringkat
- Simultaneous Determination of Methylparaben + Propylparaben + Hidrocortisone Topical Cream PDFDokumen7 halamanSimultaneous Determination of Methylparaben + Propylparaben + Hidrocortisone Topical Cream PDFNájla KassabBelum ada peringkat
- Screenshot 2564-04-16 at 09.45.04Dokumen95 halamanScreenshot 2564-04-16 at 09.45.04Sarunpong ImampaiBelum ada peringkat
- Daktari Bingwa at WorkDokumen8 halamanDaktari Bingwa at WorkMuhidin Issa MichuziBelum ada peringkat