Anda mungkin juga menyukai
- The Sound of Concepts: The Link Between Auditory and Conceptual Brain SystemsDokumen20 halamanThe Sound of Concepts: The Link Between Auditory and Conceptual Brain SystemsParagBelum ada peringkat
- Neuropsychology of Prefrontal Cortex.13Dokumen7 halamanNeuropsychology of Prefrontal Cortex.13Semma LopesBelum ada peringkat
- Neural Entrainment and Attentional SelectionDokumen14 halamanNeural Entrainment and Attentional SelectionKornél SzabóBelum ada peringkat
- Cabeza Etal 2003 NeuropsychologiaDokumen10 halamanCabeza Etal 2003 Neuropsychologiaconicet.ibymeBelum ada peringkat
- Bola Proceedingsofthe National Academyof Sciences 2017Dokumen16 halamanBola Proceedingsofthe National Academyof Sciences 2017courursulaBelum ada peringkat
- Mindfullnes and CongnitionDokumen10 halamanMindfullnes and CongnitionCamila AmorimBelum ada peringkat
- Marsh J E and Campbell T 2016 ProcessingDokumen33 halamanMarsh J E and Campbell T 2016 Processingcarlos martinezBelum ada peringkat
- Review: Musical Training As A Framework For Brain Plasticity: Behavior, Function, and StructureDokumen17 halamanReview: Musical Training As A Framework For Brain Plasticity: Behavior, Function, and Structureyokai12Belum ada peringkat
- 2013 Steinschneider Hear ResDokumen17 halaman2013 Steinschneider Hear ResseantipsBelum ada peringkat
- Repetition Suppression and Reactivation in Auditory - Verbal Short-Term Recognition MemoryDokumen12 halamanRepetition Suppression and Reactivation in Auditory - Verbal Short-Term Recognition Memoryandra_varjogheBelum ada peringkat
- Working Memory and Auditory Processing in School-Age ChildrenDokumen15 halamanWorking Memory and Auditory Processing in School-Age ChildrenvviraaBelum ada peringkat
- Mapping Cortical Hubs in Tinnitus No en La BiblioDokumen14 halamanMapping Cortical Hubs in Tinnitus No en La Biblioaitrus71Belum ada peringkat
- Jezicki MozakDokumen36 halamanJezicki MozakSinisa RisticBelum ada peringkat
- Ni Hms 858658Dokumen8 halamanNi Hms 858658Rubén Alcalá ArgüellesBelum ada peringkat
- The Brain Basis of LanguageDokumen36 halamanThe Brain Basis of LanguageGrethell UrcielBelum ada peringkat
- Recanzone Rapidly InducedDokumen7 halamanRecanzone Rapidly InducedViji ThulasiramanBelum ada peringkat
- TMP 286 ADokumen14 halamanTMP 286 AFrontiersBelum ada peringkat
- The Neural Basis of The Complex Mental Task of Meditation NewbergDokumen10 halamanThe Neural Basis of The Complex Mental Task of Meditation NewbergAlexander PriceBelum ada peringkat
- Auditory Cortical Changes Precede Brainstem Changes During Rapid Implicit Learning: Evidence From Human EEGDokumen9 halamanAuditory Cortical Changes Precede Brainstem Changes During Rapid Implicit Learning: Evidence From Human EEGCaroline RochaBelum ada peringkat
- Brain and Cognition: Ruiz-Guti Errez Jos E, Arias-S Anchez Samuel, Martín-Monz On IsabelDokumen11 halamanBrain and Cognition: Ruiz-Guti Errez Jos E, Arias-S Anchez Samuel, Martín-Monz On IsabelgustavBelum ada peringkat
- Laffere2020 - Effects of Auditory Selective Attention On Neural PhaseDokumen9 halamanLaffere2020 - Effects of Auditory Selective Attention On Neural PhaseErick SolisBelum ada peringkat
- 0956 CH13Dokumen12 halaman0956 CH13Felipe MatheusBelum ada peringkat
- Bain MechanismDokumen10 halamanBain MechanismAna VenturaBelum ada peringkat
- Flexible and Dynamic Representations of Gustatory InformationDokumen6 halamanFlexible and Dynamic Representations of Gustatory InformationDeepika GovindarajanBelum ada peringkat
- Lusk, 2016. Cerebellar Hippocampal and Striatal Time CellsDokumen7 halamanLusk, 2016. Cerebellar Hippocampal and Striatal Time CellsAnaMaríaMalagónLBelum ada peringkat
- Fnhum 08 00467Dokumen10 halamanFnhum 08 00467moiBelum ada peringkat
- Sistemul AuditivDokumen10 halamanSistemul AuditivCalugareanu MadalinaBelum ada peringkat
- Tune-Obleser SHAR 2021 FinalDokumen32 halamanTune-Obleser SHAR 2021 Finall.fernandezBelum ada peringkat
- Temporal Contrast Effects in Human Speech Perception Are Immune To Selective AttentionDokumen11 halamanTemporal Contrast Effects in Human Speech Perception Are Immune To Selective AttentionAjay PatelBelum ada peringkat
- Chapter 9 - Middle - and Long-Latency Auditory Evoked PotentialsDokumen23 halamanChapter 9 - Middle - and Long-Latency Auditory Evoked PotentialsCaio LeônidasBelum ada peringkat
- TMP 73 C3Dokumen21 halamanTMP 73 C3FrontiersBelum ada peringkat
- Neuromodulation of Prefrontal Cortex Cognitive Function in Primates - The Powerful Roles of Monoamines and AcetylcholineDokumen20 halamanNeuromodulation of Prefrontal Cortex Cognitive Function in Primates - The Powerful Roles of Monoamines and AcetylcholineDouglas DiasBelum ada peringkat
- Koelsch 2005Dokumen7 halamanKoelsch 2005freitas.pianoBelum ada peringkat
- Brain 2006 Stewart Music and BrainDokumen21 halamanBrain 2006 Stewart Music and BrainAndrada Tatiana CrișanBelum ada peringkat
- Research PaperDokumen9 halamanResearch Paperapi-282381441Belum ada peringkat
- Ambiguous Figures - What Happens in The Brain When Perception Changes But Not The StimulusDokumen23 halamanAmbiguous Figures - What Happens in The Brain When Perception Changes But Not The StimulusPriyanka ChakrabortyBelum ada peringkat
- Brain & Language: Max Garagnani, Friedemann PulvermüllerDokumen11 halamanBrain & Language: Max Garagnani, Friedemann PulvermüllerTroy CabrillasBelum ada peringkat
- Learning and Cognitive Flexibility - Frontostriatal Function and Monoaminergic ModulationDokumen6 halamanLearning and Cognitive Flexibility - Frontostriatal Function and Monoaminergic ModulationFrancisco Ahumada MéndezBelum ada peringkat
- The Memory of Noise: Christian KaernbachDokumen9 halamanThe Memory of Noise: Christian KaernbachAnnisa febriani putriBelum ada peringkat
- Normal and Pathological Oscillatory Communication in The BrainDokumen12 halamanNormal and Pathological Oscillatory Communication in The BrainDiegoBelum ada peringkat
- 01 Miodrag Stokic Zorana Milosavljevic Slavica MaksimovicDokumen12 halaman01 Miodrag Stokic Zorana Milosavljevic Slavica MaksimovicАлександра ВукосављевићBelum ada peringkat
- Functional Magnetic Resonance Imaging (fMRI) : A Window Into The BrainDokumen28 halamanFunctional Magnetic Resonance Imaging (fMRI) : A Window Into The BrainAlexia KarousouBelum ada peringkat
- The Neural Correlates of Conscious VisionDokumen14 halamanThe Neural Correlates of Conscious VisionKarla ValderramaBelum ada peringkat
- Music and The Brain StewartDokumen21 halamanMusic and The Brain StewartCatherine BoichardBelum ada peringkat
- Rausch 2014Dokumen12 halamanRausch 2014Azadi XerxesBelum ada peringkat
- The Prefronal Cortex and Flexible BehaviorDokumen25 halamanThe Prefronal Cortex and Flexible BehaviorrorozainosBelum ada peringkat
- Plasticity, Auditory Training, and Auditory Processing DisordersDokumen14 halamanPlasticity, Auditory Training, and Auditory Processing DisordersLuis SeixasBelum ada peringkat
- Nihms235910 PDFDokumen10 halamanNihms235910 PDFgenerjustnBelum ada peringkat
- The Modern Concept of Association Cortex Vanhoesen1993Dokumen5 halamanThe Modern Concept of Association Cortex Vanhoesen1993LucíaBelum ada peringkat
- Working Memory FMRIDokumen9 halamanWorking Memory FMRIViktor NybergBelum ada peringkat
- The Role of Left Inferior Frontal and Superior Temporal Cortex in Sentence Comprehension: Localizing Syntactic and Semantic ProcessesDokumen8 halamanThe Role of Left Inferior Frontal and Superior Temporal Cortex in Sentence Comprehension: Localizing Syntactic and Semantic ProcessesBruno PerazioBelum ada peringkat
- Buzsaki 2004 Nat NeurosciDokumen7 halamanBuzsaki 2004 Nat Neuroscihussin elrashidyBelum ada peringkat
- Whitford, 2019 - ReviewDokumen14 halamanWhitford, 2019 - ReviewRosa BeñoBelum ada peringkat
- 2004 Towards Studies of The Social BrainDokumen5 halaman2004 Towards Studies of The Social BrainAl WilsonBelum ada peringkat
- ShowDokumen123 halamanShowStarLink1Belum ada peringkat
- Stress Signalling Pathways and Prefrontal FuctionsDokumen13 halamanStress Signalling Pathways and Prefrontal FuctionsFrancisco MtzBelum ada peringkat
- Representing Inner Voices in VRDokumen10 halamanRepresenting Inner Voices in VRCaio Cesar LouresBelum ada peringkat
- Neuroimagen y EmocionDokumen10 halamanNeuroimagen y Emocionnatsumi18Belum ada peringkat
- Understanding Cognitive Neuroscience: A Simplified GuideDari EverandUnderstanding Cognitive Neuroscience: A Simplified GuideBelum ada peringkat
- Psych9223 F2014 L08 GroupDataDokumen67 halamanPsych9223 F2014 L08 GroupDataCassia LmtBelum ada peringkat
- fMRI Data Quality Assurance and PreprocessingDokumen84 halamanfMRI Data Quality Assurance and PreprocessingCassia LmtBelum ada peringkat
- fMRI Physics in A Nutshell: Understanding WTF Your MR Physicist Is Talking AboutDokumen72 halamanfMRI Physics in A Nutshell: Understanding WTF Your MR Physicist Is Talking AboutCassia LmtBelum ada peringkat
- fMRI Data Quality Assurance and PreprocessingDokumen84 halamanfMRI Data Quality Assurance and PreprocessingCassia LmtBelum ada peringkat
- Basics of Experimental Design For fMRI: Event-Related DesignsDokumen101 halamanBasics of Experimental Design For fMRI: Event-Related DesignsCassia LmtBelum ada peringkat
- fMRI Analysis with the General Linear ModelDokumen121 halamanfMRI Analysis with the General Linear ModelCassia LmtBelum ada peringkat
- Basics of Experimental Design For fMRI: Block DesignsDokumen99 halamanBasics of Experimental Design For fMRI: Block DesignsCassia LmtBelum ada peringkat
- AnsysDokumen26 halamanAnsysMike BillBelum ada peringkat
- RiceX - Discrete Time Signals and Systems, Part 2 Freq Domain WK 2Dokumen2 halamanRiceX - Discrete Time Signals and Systems, Part 2 Freq Domain WK 2Cassia LmtBelum ada peringkat
- ELEC301.1x Discrete Time Signals and Systems, Part 2: Frequency Domain (brief course notesDokumen5 halamanELEC301.1x Discrete Time Signals and Systems, Part 2: Frequency Domain (brief course notesCassia LmtBelum ada peringkat
- Psych9223 F2014 L08 GroupDataDokumen67 halamanPsych9223 F2014 L08 GroupDataCassia LmtBelum ada peringkat
- RiceX - Discrete Time Signals and Systems, Part 1 Time DomainDokumen5 halamanRiceX - Discrete Time Signals and Systems, Part 1 Time DomainCassia LmtBelum ada peringkat
- DTFT Analysis of Infinite SignalsDokumen50 halamanDTFT Analysis of Infinite SignalsCassia LmtBelum ada peringkat
- Basics of Experimental Design For fMRI: Block DesignsDokumen99 halamanBasics of Experimental Design For fMRI: Block DesignsCassia LmtBelum ada peringkat
- Online CoursesDokumen2 halamanOnline CoursesCassia LmtBelum ada peringkat
- ELEC301 2x-Week1 Rev2Dokumen93 halamanELEC301 2x-Week1 Rev2Cassia LmtBelum ada peringkat
- Heroes3 Manual AB PDFDokumen29 halamanHeroes3 Manual AB PDFNicolas ManettaBelum ada peringkat
- HOMM3Dokumen144 halamanHOMM3Nistor Florin100% (1)
- Brains Bodies Behaviour (Psychology)Dokumen18 halamanBrains Bodies Behaviour (Psychology)Cassia LmtBelum ada peringkat
- CH 7Dokumen17 halamanCH 7Ginny MontalbanBelum ada peringkat
- Controlling The False Discovery RateDokumen13 halamanControlling The False Discovery RateCassia LmtBelum ada peringkat
- MEGDokumen1 halamanMEGCassia LmtBelum ada peringkat
- Lesson 2 - Applications of Valuing Cash FlowsDokumen10 halamanLesson 2 - Applications of Valuing Cash FlowsStephen MagudhaBelum ada peringkat
- Memory (Psychology)Dokumen25 halamanMemory (Psychology)Cassia LmtBelum ada peringkat
- Statistics Basics, Distributions, Confidence Intervals, Hypothesis TestingDokumen3 halamanStatistics Basics, Distributions, Confidence Intervals, Hypothesis TestingCassia LmtBelum ada peringkat
- Imperativopasado - irregularesBSP103x Courseware - EdxDokumen3 halamanImperativopasado - irregularesBSP103x Courseware - EdxCassia LmtBelum ada peringkat
- H3SoD PC ManualDokumen36 halamanH3SoD PC ManualvitorclaudinoBelum ada peringkat
- Heroes III TutorialDokumen12 halamanHeroes III TutorialzaralaviniaBelum ada peringkat
- Valuing CashflowsDokumen11 halamanValuing CashflowsAbelBelum ada peringkat
- HOMM3Dokumen144 halamanHOMM3Nistor Florin100% (1)
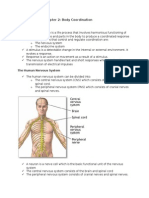
- Form 4 Science Chapter 2: Body Coordination (38Dokumen13 halamanForm 4 Science Chapter 2: Body Coordination (38Cheah Foo Kit50% (2)
- The Evolutionary Basis of Strabismus and Nystagmus in ChildrenDokumen244 halamanThe Evolutionary Basis of Strabismus and Nystagmus in ChildrenComan MihaelaBelum ada peringkat
- Basal Ganglia Circuits As Targets For Neuromodulation in Parkinson DiseaseDokumen7 halamanBasal Ganglia Circuits As Targets For Neuromodulation in Parkinson DiseaseSebastián GallegosBelum ada peringkat
- Paxinos The Rat Brain 73Dokumen1 halamanPaxinos The Rat Brain 73joseuri2011Belum ada peringkat
- Clinical Examination of Patients With Headache ComplaintsDokumen157 halamanClinical Examination of Patients With Headache ComplaintsrapannikaBelum ada peringkat
- COMITANT CONVERGENT STRABISMUS WITH ACUTE ONSETDokumen10 halamanCOMITANT CONVERGENT STRABISMUS WITH ACUTE ONSETAgung Bhakti WiratamaBelum ada peringkat
- Final Study Guide Psyc101 Spring 2011Dokumen14 halamanFinal Study Guide Psyc101 Spring 2011FirstLady Williams100% (1)
- Anatomy & Physiology of Olfactory System.Dokumen27 halamanAnatomy & Physiology of Olfactory System.Prasanna DattaBelum ada peringkat
- Mri Brain SequencesDokumen13 halamanMri Brain SequencesRisa MarissaBelum ada peringkat
- Psychiatry: Taking A Psychiatric HistoryDokumen48 halamanPsychiatry: Taking A Psychiatric Historyfdjslajf100% (1)
- Yokukansan (TJ-54) in The Treatment Neurological Disorders - A ReviewDokumen6 halamanYokukansan (TJ-54) in The Treatment Neurological Disorders - A ReviewAurelianBelum ada peringkat
- SchizophreniaDokumen34 halamanSchizophreniaSarasija NatarajanBelum ada peringkat
- Hebb 1949Dokumen16 halamanHebb 1949Norel Nicolae BalutaBelum ada peringkat
- Osama - S Ophthalmology Notes FEBRUARY 2019 EDITIONDokumen42 halamanOsama - S Ophthalmology Notes FEBRUARY 2019 EDITIONNazish Saghir67% (3)
- Principles of Neural CircuitsDokumen6 halamanPrinciples of Neural CircuitsMarius MyburgBelum ada peringkat
- Cervical Spondylosis: Definition, Epidemiology, Clinical PresentationDokumen14 halamanCervical Spondylosis: Definition, Epidemiology, Clinical PresentationJaspreet kaurBelum ada peringkat
- Spatial Hearing: The Psychophysics of Human Sound LocalizationDokumen3 halamanSpatial Hearing: The Psychophysics of Human Sound LocalizationDaniel Ramos FernándezBelum ada peringkat
- Several" and Graph Which Means "Writings." Hence, Literally, Polygraph Means ManyDokumen34 halamanSeveral" and Graph Which Means "Writings." Hence, Literally, Polygraph Means Manykim ryan uchiBelum ada peringkat
- Amyotrophic Lateral Sclerosis: PathophysiologyDokumen4 halamanAmyotrophic Lateral Sclerosis: PathophysiologyJulia SalvioBelum ada peringkat
- Ocular and Visual Physiology - Clinical ApplicationDokumen372 halamanOcular and Visual Physiology - Clinical ApplicationGrecia Sánchez RuízBelum ada peringkat
- Discontinuation Discharge NoteDokumen3 halamanDiscontinuation Discharge Noteapi-488383880Belum ada peringkat
- CH 5 ElectricityDokumen39 halamanCH 5 ElectricityAbdullahi Mohamed IsakBelum ada peringkat
- Innocent Lamola Department of Health & Temperance Mowbray SDA Church & SDASM UCTDokumen15 halamanInnocent Lamola Department of Health & Temperance Mowbray SDA Church & SDASM UCTshuaibgogaBelum ada peringkat
- Functional NeuroanatomyDokumen23 halamanFunctional NeuroanatomyDaniela GrigoraşBelum ada peringkat
- The Ventricular SystemDokumen6 halamanThe Ventricular SystemM Arsalan TariqBelum ada peringkat
- Marsha Linehan - A biosocial developmental model of borderline personality: Elaborating and extending Linehan's theoryDokumen16 halamanMarsha Linehan - A biosocial developmental model of borderline personality: Elaborating and extending Linehan's theoryIrving Pérez MéndezBelum ada peringkat
- Chapter 3 PretestDokumen18 halamanChapter 3 PretestAl GilaniBelum ada peringkat
- World Through Our Senses (All Sensory Organs)Dokumen48 halamanWorld Through Our Senses (All Sensory Organs)Chin Kok Soon100% (3)
- Biology Notes 12 Sem2Dokumen11 halamanBiology Notes 12 Sem2josmanlilyBelum ada peringkat
- A Study On Triplet Repeat Expansion Disorders in Western Indian PopulationDokumen5 halamanA Study On Triplet Repeat Expansion Disorders in Western Indian PopulationShavana RajkumarBelum ada peringkat