
Anda mungkin juga menyukai
- Atm 04 08 160Dokumen9 halamanAtm 04 08 160manalBelum ada peringkat
- 178 PDFDokumen6 halaman178 PDFmanalBelum ada peringkat
- A Focus On Respiratory Tractinfection As An Important Risk Factor in CopdDokumen3 halamanA Focus On Respiratory Tractinfection As An Important Risk Factor in CopdmanalBelum ada peringkat
- Ajr.13.11288 FixDokumen12 halamanAjr.13.11288 FixmanalBelum ada peringkat
- DRP2012 303275Dokumen6 halamanDRP2012 303275PoncoBelum ada peringkat
- 6 The Use of Controller Medications PDFDokumen7 halaman6 The Use of Controller Medications PDFmanalBelum ada peringkat
- 2016 Article 1334Dokumen12 halaman2016 Article 1334manalBelum ada peringkat
- Ves - 2015 - 25 3,4 - Ves 25 3,4 Ves553 - Ves 25 Ves553Dokumen13 halamanVes - 2015 - 25 3,4 - Ves 25 3,4 Ves553 - Ves 25 Ves553Nadira Juanti PratiwiBelum ada peringkat
- 1 s2.0 S0975947616301462 Main PDFDokumen6 halaman1 s2.0 S0975947616301462 Main PDFmanalBelum ada peringkat
- Association of BMI, socioeconomic status and menarche age with anemiaDokumen7 halamanAssociation of BMI, socioeconomic status and menarche age with anemiamanalBelum ada peringkat
- Effect of Microcytic Hypochromic Anemia and Parasitic Infestations OnDokumen9 halamanEffect of Microcytic Hypochromic Anemia and Parasitic Infestations OnmanalBelum ada peringkat
- InTech-Nutritional Anemia in Developing CountriesDokumen21 halamanInTech-Nutritional Anemia in Developing CountriesmanalBelum ada peringkat
- Jagaa YooDokumen1 halamanJagaa YoomanalBelum ada peringkat
- Iron Deficiency and Iron Deficiency Anemia in AdolescenDokumen1 halamanIron Deficiency and Iron Deficiency Anemia in AdolescenmanalBelum ada peringkat
- Dietary Pattern and Factors Associated With Anemia AmongDokumen11 halamanDietary Pattern and Factors Associated With Anemia AmongmanalBelum ada peringkat
- 409 Ijar-5331Dokumen6 halaman409 Ijar-5331Nia UtariBelum ada peringkat
- A Novel Technique For The Treatment of Stages III.46 PDFDokumen5 halamanA Novel Technique For The Treatment of Stages III.46 PDFmanalBelum ada peringkat
- Anes Mnge Burn 1212 p430 439 PDFDokumen10 halamanAnes Mnge Burn 1212 p430 439 PDFmanalBelum ada peringkat
- DAFTAR PUSTAKA 28 JULI 2015.docx - 0Dokumen3 halamanDAFTAR PUSTAKA 28 JULI 2015.docx - 0manalBelum ada peringkat
- Ipi200728 PDFDokumen14 halamanIpi200728 PDFNovi LatifaBelum ada peringkat
- ReferenceDokumen8 halamanReferencePutri YunianingsihBelum ada peringkat
- GNL 1123Dokumen9 halamanGNL 1123manalBelum ada peringkat
- Tard 43 3 154 161 (A)Dokumen8 halamanTard 43 3 154 161 (A)manalBelum ada peringkat
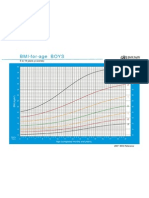
- Bmi For Age Z-ScoreDokumen1 halamanBmi For Age Z-ScoreTisha Patricia OedoyBelum ada peringkat
- Prevalensi Candidiasis 2013Dokumen9 halamanPrevalensi Candidiasis 2013manalBelum ada peringkat
- Cold Burn Injuries in The UK: The 11-Year Experience of A Tertiary Burns CentreDokumen8 halamanCold Burn Injuries in The UK: The 11-Year Experience of A Tertiary Burns CentremanalBelum ada peringkat
- Stress Ulcer Versus PlaceboDokumen8 halamanStress Ulcer Versus PlacebomanalBelum ada peringkat
- Tard 43 3 154 161 (A)Dokumen8 halamanTard 43 3 154 161 (A)manalBelum ada peringkat
- Congenital UrogenitalDokumen6 halamanCongenital UrogenitalmanalBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5784)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (72)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- BES-121 Childhood and Growing UpDokumen57 halamanBES-121 Childhood and Growing UpPulkesh PulakBelum ada peringkat
- Mr. Russell and The Reconstruction of Society Essay by AmbedkarDokumen5 halamanMr. Russell and The Reconstruction of Society Essay by AmbedkarthakurrrBelum ada peringkat
- TheoryDokumen5 halamanTheoryCaro-lene Dela CruzBelum ada peringkat
- BSP and GSP ProposalDokumen7 halamanBSP and GSP ProposalHylord Guzman100% (2)
- Effects of Instructional Materials On The Teaching and Learning of Social Studies in Secondary Schools in Oju Local Government Area of Benue StateDokumen6 halamanEffects of Instructional Materials On The Teaching and Learning of Social Studies in Secondary Schools in Oju Local Government Area of Benue StateSamjay wantedBelum ada peringkat
- Facial Expression Recognition CNN SurveyDokumen4 halamanFacial Expression Recognition CNN Surveypapunjay kumarBelum ada peringkat
- SCAS Girls 8-11 FinalJune2018Dokumen1 halamanSCAS Girls 8-11 FinalJune2018Gonzalo GVBelum ada peringkat
- Urban Entertainment DestinationsDokumen12 halamanUrban Entertainment DestinationsBilal Yousaf50% (2)
- A Project Manager's OptimismDokumen18 halamanA Project Manager's OptimismIna NafaBelum ada peringkat
- Written Report in Eed Eng 2Dokumen4 halamanWritten Report in Eed Eng 2paula macatangayBelum ada peringkat
- Example Interview QuestionsDokumen3 halamanExample Interview Questionsapi-3745830Belum ada peringkat
- Final Essay - Signature AssignmentDokumen6 halamanFinal Essay - Signature Assignmentapi-600669880Belum ada peringkat
- GL Bajaj FormateDokumen5 halamanGL Bajaj FormateFARMANBelum ada peringkat
- Unit 3.3 Cognitive Processes: Constructivism: Knowledge Construction/ LearningDokumen5 halamanUnit 3.3 Cognitive Processes: Constructivism: Knowledge Construction/ LearningCherry Rose J. Deniega100% (1)
- nstp1 Final ExamDokumen2 halamannstp1 Final ExamRichevin Denzo100% (1)
- Cultural Traits Can Assume Various Forms Which Have Meanings. The Statement Implies That Culture IsDokumen21 halamanCultural Traits Can Assume Various Forms Which Have Meanings. The Statement Implies That Culture IsmercyvienhoBelum ada peringkat
- Organizational DevelopmentDokumen29 halamanOrganizational DevelopmentvaijayanthiBelum ada peringkat
- Ispo Self-Study DocumentDokumen24 halamanIspo Self-Study DocumentFatima SanchezBelum ada peringkat
- Application LetterDokumen1 halamanApplication LetterEvan Carlos MajaduconBelum ada peringkat
- Types of Training and Its BenefitsDokumen12 halamanTypes of Training and Its BenefitsNajmiBelum ada peringkat
- Hexagram 41, Decreasing: Howshudimoveforward01122019Dokumen3 halamanHexagram 41, Decreasing: Howshudimoveforward01122019rotBelum ada peringkat
- Modern Approaches and Innovation in PsychologyDokumen206 halamanModern Approaches and Innovation in Psychologykohinoorncmaip67% (3)
- EthicsDokumen6 halamanEthicsCess UrrizaBelum ada peringkat
- The Dangers of GroupthinkDokumen14 halamanThe Dangers of GroupthinkSushil Aggrawal100% (1)
- Defensive coping nursing diagnosisDokumen3 halamanDefensive coping nursing diagnosisRoch Oconer100% (1)
- Reading and Writing 3 Q: Skills For Success Unit 6 Student Book Answer KeyDokumen3 halamanReading and Writing 3 Q: Skills For Success Unit 6 Student Book Answer Keybichhanh142Belum ada peringkat
- Japan Doing Business in a Unique Culture (Bucknall, Kevin B. [Bucknall, Kevin B.]) (Z-Library)Dokumen218 halamanJapan Doing Business in a Unique Culture (Bucknall, Kevin B. [Bucknall, Kevin B.]) (Z-Library)sparta151122Belum ada peringkat
- Principios de Circuitos ElectricosOKDokumen121 halamanPrincipios de Circuitos ElectricosOKerechBelum ada peringkat
- Boston Qualitative Scoring System for Rey-Osterrieth Complex Figure Effective for Detecting Cognitive Impairment in Parkinson's DiseaseDokumen9 halamanBoston Qualitative Scoring System for Rey-Osterrieth Complex Figure Effective for Detecting Cognitive Impairment in Parkinson's DiseaseJuanBelum ada peringkat
- Assessment in SchoolDokumen3 halamanAssessment in SchoolCel Rellores SalazarBelum ada peringkat





























































































![Japan Doing Business in a Unique Culture (Bucknall, Kevin B. [Bucknall, Kevin B.]) (Z-Library)](https://imgv2-2-f.scribdassets.com/img/document/721402476/149x198/886c54a86e/1712650374?v=1)


