Anda mungkin juga menyukai
- Eizenberg S General Anatomy PDFDokumen255 halamanEizenberg S General Anatomy PDFNikita Leanne FernandesBelum ada peringkat
- Descriptive Words and PhrasesDokumen39 halamanDescriptive Words and PhrasesShukri Ayob100% (1)
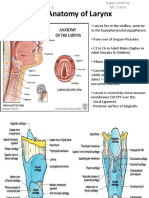
- Anatomy of Larynx - Wai Sheng Xuan Batch 3 1101A11688Dokumen27 halamanAnatomy of Larynx - Wai Sheng Xuan Batch 3 1101A11688Roshan Gill100% (1)
- Brain Teasers 2Dokumen2 halamanBrain Teasers 2Starpiter50% (6)
- Segregating Variation For Temperature-Dependent Sex Determination in A LizardDokumen12 halamanSegregating Variation For Temperature-Dependent Sex Determination in A LizardUrielBelum ada peringkat
- Evaluation of Conception Rate in Nordic Dairy CattleDokumen6 halamanEvaluation of Conception Rate in Nordic Dairy CattleH-diBelum ada peringkat
- 0141 (1) EeDokumen4 halaman0141 (1) EeMariana MansillaBelum ada peringkat
- A Field Study On Fertility and Purity of Sex-Sorted Cattle SpermDokumen5 halamanA Field Study On Fertility and Purity of Sex-Sorted Cattle Spermyara salesBelum ada peringkat
- Paternally Induced Transgenerational Environmental Reprogramming of Metabolic Gene Expression in MammalsDokumen13 halamanPaternally Induced Transgenerational Environmental Reprogramming of Metabolic Gene Expression in MammalsAshok KumarBelum ada peringkat
- Allelic and Population Variation of Microsatellite Loci in Aspen (Populus Tremuloides)Dokumen10 halamanAllelic and Population Variation of Microsatellite Loci in Aspen (Populus Tremuloides)Anny LovibBelum ada peringkat
- Molecular Support For Heterogonesis Resulting in Sesquizygotic TwinningDokumen8 halamanMolecular Support For Heterogonesis Resulting in Sesquizygotic TwinningartBelum ada peringkat
- s41598 020 67089 0 PDFDokumen11 halamans41598 020 67089 0 PDFErick Saldaña VillaBelum ada peringkat
- Association Between Single Nucleotide Polymorphisms Andsexual Precocity in Nellore HeifersDokumen9 halamanAssociation Between Single Nucleotide Polymorphisms Andsexual Precocity in Nellore HeifersInaê RegatieriBelum ada peringkat
- Artikel Implant 1Dokumen9 halamanArtikel Implant 1Much MahmudiBelum ada peringkat
- Inbreeding: Biotechnology in Animal Husbandry January 2007Dokumen19 halamanInbreeding: Biotechnology in Animal Husbandry January 2007Harry ChristamaBelum ada peringkat
- Concept of Genomic SelectionDokumen35 halamanConcept of Genomic SelectionKush ShrivastavaBelum ada peringkat
- 01.BAH 19 4 9 29.11.2019. Petrović M. P. Et Al.Dokumen11 halaman01.BAH 19 4 9 29.11.2019. Petrović M. P. Et Al.Ivan PavlovicBelum ada peringkat
- Research Article: Jin Jun, Paul Ryvkin, Edward Hemphill, Ion Mandoiu, and Craig NelsonDokumen16 halamanResearch Article: Jin Jun, Paul Ryvkin, Edward Hemphill, Ion Mandoiu, and Craig Nelsonzune153Belum ada peringkat
- BROWN Et Al-2007-Journal of Evolutionary BiologyDokumen9 halamanBROWN Et Al-2007-Journal of Evolutionary BiologyPrabodhini Upadhyay100% (1)
- Katona Et Al. 2021 JEBDokumen10 halamanKatona Et Al. 2021 JEBSomogyvári PetraBelum ada peringkat
- The Phylogeny of Staphylococcus Aureus - Which Genes Make The Best Intra-Species Markers?Dokumen9 halamanThe Phylogeny of Staphylococcus Aureus - Which Genes Make The Best Intra-Species Markers?lukman hakimBelum ada peringkat
- Animal Breeding MethodsDokumen186 halamanAnimal Breeding MethodsAmanuel AsratBelum ada peringkat
- Heritability of Sudden Death Syndrome and Its Associated Correlations To Ascites and Body Weight in BroilersDokumen4 halamanHeritability of Sudden Death Syndrome and Its Associated Correlations To Ascites and Body Weight in BroilersSuraj_SubediBelum ada peringkat
- An Autosomal Genomic Screen For Autism 1999Dokumen7 halamanAn Autosomal Genomic Screen For Autism 1999Harshit AmbeshBelum ada peringkat
- Buletin PeternakanDokumen5 halamanBuletin PeternakanLatifah LymafBelum ada peringkat
- PIIS0022030213000568Dokumen10 halamanPIIS0022030213000568Laura PîndaruBelum ada peringkat
- Reproductive Failure 2Dokumen8 halamanReproductive Failure 2apoyo7royal park parkBelum ada peringkat
- Sweden Twins Same SexDokumen6 halamanSweden Twins Same SexmarcellojunBelum ada peringkat
- 10 3168@jds 2009-2720 en IdDokumen14 halaman10 3168@jds 2009-2720 en IdUswatun HasanahBelum ada peringkat
- Kaiser Et Al. 2016 Mer PDFDokumen11 halamanKaiser Et Al. 2016 Mer PDFAditya WirawanBelum ada peringkat
- Cáceres 2009Dokumen14 halamanCáceres 2009Manu Gustani BussBelum ada peringkat
- Genetic Diversity of Dog Breeds Within-Breed Diversity Comparing Genealogical and Molecular DataDokumen10 halamanGenetic Diversity of Dog Breeds Within-Breed Diversity Comparing Genealogical and Molecular DatakadaBelum ada peringkat
- Genetic Control of Reproduction in Dairy Cows: Stephen T. ButlerDokumen11 halamanGenetic Control of Reproduction in Dairy Cows: Stephen T. ButlerAndres LeonBelum ada peringkat
- A Description of The Growth of Sheep and Its Genetic AnalysisDokumen13 halamanA Description of The Growth of Sheep and Its Genetic AnalysisYessenia FigueroaBelum ada peringkat
- Author's Accepted Manuscript: Livestock ScienceDokumen15 halamanAuthor's Accepted Manuscript: Livestock ScienceDomínguez Derik EriksenkBelum ada peringkat
- Mark Scheme 2015Dokumen6 halamanMark Scheme 2015api-324166624Belum ada peringkat
- 1 s2.0 S0022030211005364 MainDokumen16 halaman1 s2.0 S0022030211005364 MainAlejandra SánchezBelum ada peringkat
- Psiki 2Dokumen7 halamanPsiki 2Winona ProkBelum ada peringkat
- Male Female RatioDokumen4 halamanMale Female RatiomaravindanBelum ada peringkat
- Jasienska Et Al-2006-American Journal of Human BiologyDokumen4 halamanJasienska Et Al-2006-American Journal of Human BiologyChristiAn OmArBelum ada peringkat
- A Mechanism For Impaired Fear Recognition After Amygdala Damage - Adolphs Et Al - 2005Dokumen5 halamanA Mechanism For Impaired Fear Recognition After Amygdala Damage - Adolphs Et Al - 2005ekitani6817Belum ada peringkat
- Evidence For Large Domains of Similarly ExpressedDokumen3 halamanEvidence For Large Domains of Similarly ExpressedKookie JunBelum ada peringkat
- Lovell 2014 MolEco Mating System and Environmental Variation Drive Patterns of Adapt in B SpatifoliaDokumen12 halamanLovell 2014 MolEco Mating System and Environmental Variation Drive Patterns of Adapt in B SpatifoliaAJMRBelum ada peringkat
- The Ashbya Gossypii Genome As A Tool For Mapping The AncientDokumen4 halamanThe Ashbya Gossypii Genome As A Tool For Mapping The AncientJuan RamirezBelum ada peringkat
- Genome Editing Livestock Breeding ProgramsDokumen17 halamanGenome Editing Livestock Breeding Programsmirielys.rondaBelum ada peringkat
- Principle of Animal BreedingDokumen7 halamanPrinciple of Animal BreedingFerlan C. Tagdulang100% (1)
- Genetic DiversityDokumen55 halamanGenetic DiversityMohammad IbrarBelum ada peringkat
- Nature 04559Dokumen4 halamanNature 04559Benjan't PujadasBelum ada peringkat
- 2021-Rates of Contributory de Novo Mutation in High and Low-Risk Autism Families.Dokumen10 halaman2021-Rates of Contributory de Novo Mutation in High and Low-Risk Autism Families.熊佳仪(不冬眠的熊)Belum ada peringkat
- Color Cáscara y EdadDokumen8 halamanColor Cáscara y Edadjorge_bustos_1Belum ada peringkat
- Comparative Transcriptome Analysis Among Parental Inbred and Crosses Reveals The Role of Dominance Gene Expression in Heterosis in DrosophilaDokumen7 halamanComparative Transcriptome Analysis Among Parental Inbred and Crosses Reveals The Role of Dominance Gene Expression in Heterosis in Drosophila廖晨延Belum ada peringkat
- Penetraciion Espermatica en AlpacasDokumen9 halamanPenetraciion Espermatica en AlpacasDaniel Gandarillas EspezuaBelum ada peringkat
- Genetic Diversity of Snps Associated With Candidate Genes For Heat Stress in Coreño Creole Cattle in MexicoDokumen7 halamanGenetic Diversity of Snps Associated With Candidate Genes For Heat Stress in Coreño Creole Cattle in Mexicorodriguez.carmen6402Belum ada peringkat
- Pnas00101 0183 PDFDokumen4 halamanPnas00101 0183 PDFGAMEPLAY & FUNNY MOMENTSBelum ada peringkat
- Biological Identifications of Mayflies (Ephemeroptera) Using DNA BarcodesDokumen17 halamanBiological Identifications of Mayflies (Ephemeroptera) Using DNA BarcodesAdirfan PratomoBelum ada peringkat
- Albatross Young Et Al.Dokumen3 halamanAlbatross Young Et Al.giuliaBelum ada peringkat
- By Alasdair Simpson User On 03 May 2018Dokumen3 halamanBy Alasdair Simpson User On 03 May 2018Gustav MolMedBelum ada peringkat
- Genomewide Scan of Hoarding in Sib Pairs in Which Both Sibs Have Gilles de La Tourette SyndromeDokumen9 halamanGenomewide Scan of Hoarding in Sib Pairs in Which Both Sibs Have Gilles de La Tourette SyndromeCita Tri KusumaBelum ada peringkat
- Schanze 2011Dokumen6 halamanSchanze 2011Fahrunnisa NurdinBelum ada peringkat
- Effect of Age and Parity On Litter Size and OffsprDokumen7 halamanEffect of Age and Parity On Litter Size and OffsprAulia Fauziyyah RahmiBelum ada peringkat
- Article AboutDokumen8 halamanArticle AboutJeremy JoyceBelum ada peringkat
- 1312 FullDokumen7 halaman1312 FullJimmyboyBelum ada peringkat
- Dawson 1997Dokumen8 halamanDawson 1997Facundo BinelliBelum ada peringkat
- Evolutionary processes and theoryDari EverandEvolutionary processes and theoryEvitar NevoBelum ada peringkat
- R265 FullDokumen13 halamanR265 FullMuch MahmudiBelum ada peringkat
- Artikel Implant 1Dokumen9 halamanArtikel Implant 1Much MahmudiBelum ada peringkat
- Client's Name Account No. Address AE Code Date: AttentionDokumen6 halamanClient's Name Account No. Address AE Code Date: AttentionMuch MahmudiBelum ada peringkat
- Tanggalopenhighlowclose H - O O - L Pivot Points ACT Hasil: Pergerakan Harga Emas Tahun 2011Dokumen65 halamanTanggalopenhighlowclose H - O O - L Pivot Points ACT Hasil: Pergerakan Harga Emas Tahun 2011Much MahmudiBelum ada peringkat
- Daftar CustomerDokumen46 halamanDaftar CustomerMuch Mahmudi100% (1)
- Histo (MCQ) PDFDokumen6 halamanHisto (MCQ) PDFstuffBelum ada peringkat
- Test Your Memory: Good Citizens Always Wear Stout ShoesDokumen3 halamanTest Your Memory: Good Citizens Always Wear Stout ShoesdanielaBelum ada peringkat
- Position Paper ZABATEDokumen20 halamanPosition Paper ZABATEAtty. Emmanuel SandichoBelum ada peringkat
- Strength Treatise by C A Sampson PDFDokumen250 halamanStrength Treatise by C A Sampson PDFJack100% (1)
- The Pittston Dispatch 06-10-2012Dokumen70 halamanThe Pittston Dispatch 06-10-2012The Times LeaderBelum ada peringkat
- BG QuestsDokumen71 halamanBG QuestsJeffrey DamherBelum ada peringkat
- Comparative and Superlative Exercises - CuencaDokumen9 halamanComparative and Superlative Exercises - CuencaNinoska CuencaBelum ada peringkat
- Ingles 5º Unit 3 PDFDokumen5 halamanIngles 5º Unit 3 PDFMiguel Angel Llamas DelicadoBelum ada peringkat
- Dog Meat ThesisDokumen10 halamanDog Meat Thesisaldrinrimorin3Belum ada peringkat
- Map & Tree Map.: Figure 1: Circle Map (Sources Maps - HTML)Dokumen8 halamanMap & Tree Map.: Figure 1: Circle Map (Sources Maps - HTML)Hus HusainiBelum ada peringkat
- Cloze Test VocabDokumen15 halamanCloze Test VocabSurachai Rodngam100% (1)
- Rembrandt Corpus BueyDokumen24 halamanRembrandt Corpus BueyManuel100% (1)
- 100 Years of ZsiDokumen5 halaman100 Years of ZsiAshutoshkumar jhaBelum ada peringkat
- Project 4ed - Level 2 - Photocopiable Activities - AudioscriptsDokumen4 halamanProject 4ed - Level 2 - Photocopiable Activities - AudioscriptsSzilvi KonczolBelum ada peringkat
- Infective Endocarditis Case ReportDokumen40 halamanInfective Endocarditis Case Reportliu_owen17100% (1)
- Laser Enhancement ProtocolsDokumen25 halamanLaser Enhancement ProtocolsWolfManBelum ada peringkat
- 34 English Spelling TodayDokumen13 halaman34 English Spelling TodayabdulmusaverBelum ada peringkat
- Singaporian Math 1Dokumen16 halamanSingaporian Math 1nimanima50Belum ada peringkat
- Urinary System-1Dokumen48 halamanUrinary System-1Muhammad SajjadBelum ada peringkat
- Blood Analyses and Anatomy of The Atlantic Salmon: Salmo SalarDokumen14 halamanBlood Analyses and Anatomy of The Atlantic Salmon: Salmo SalarSenay MehariBelum ada peringkat
- PPG Reviewer For ShsDokumen5 halamanPPG Reviewer For ShsGoldwin RamirezBelum ada peringkat
- Type I HypersensitivityDokumen2 halamanType I HypersensitivitySteven MatualiBelum ada peringkat
- De Waal, F. 2014. The Bonobo and The Atheist. in Search of Humanism Among The Primates PDFDokumen4 halamanDe Waal, F. 2014. The Bonobo and The Atheist. in Search of Humanism Among The Primates PDFMelchiorBelum ada peringkat
- A Hebrew Primer BallDokumen288 halamanA Hebrew Primer Ballondoveo4749Belum ada peringkat
- Noun Proper and CommonDokumen13 halamanNoun Proper and CommonSurya BarrytappyBelum ada peringkat
- Pemeriksaan Anemia SMP Darul MaarifDokumen38 halamanPemeriksaan Anemia SMP Darul MaarifPuskesmas BanyuputihBelum ada peringkat