Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Pressure Drop (MM H2O) Water Flowrate 0 L/min 2 L/min 3 L/min 4 L/min Air FlowrateDokumen11 halamanPressure Drop (MM H2O) Water Flowrate 0 L/min 2 L/min 3 L/min 4 L/min Air FlowrateMelchiBelum ada peringkat
- 2.01 Surface Kinetics - Unimolecular Reactions PDFDokumen33 halaman2.01 Surface Kinetics - Unimolecular Reactions PDFMelchiBelum ada peringkat
- Integration of The Michaelis-Menten EquationDokumen2 halamanIntegration of The Michaelis-Menten EquationMelchiBelum ada peringkat
- Preparation of Papers For IEEE T and J: (December 2013)Dokumen8 halamanPreparation of Papers For IEEE T and J: (December 2013)MelchiBelum ada peringkat
- Water Financial Report BasicDokumen2 halamanWater Financial Report BasicMelchiBelum ada peringkat
- I. Introduction & A Brief HistoryDokumen2 halamanI. Introduction & A Brief HistoryMelchiBelum ada peringkat
- Experiment 4 - Quantitative Analysis of Soda Ash by Double-Indicator TitrationDokumen2 halamanExperiment 4 - Quantitative Analysis of Soda Ash by Double-Indicator TitrationMelchiBelum ada peringkat
- L15 SemiconductorsDiodes 1Dokumen26 halamanL15 SemiconductorsDiodes 1MelchiBelum ada peringkat
- 101.4 Mass Balance 2Dokumen10 halaman101.4 Mass Balance 2Melchi100% (2)
- Damgo ATQ Ex.5Dokumen2 halamanDamgo ATQ Ex.5MelchiBelum ada peringkat
- Damgo ATQ Ex.5Dokumen2 halamanDamgo ATQ Ex.5MelchiBelum ada peringkat
- Physics 72.1 Ex2Dokumen4 halamanPhysics 72.1 Ex2MelchiBelum ada peringkat
- Data SheetDokumen14 halamanData SheetMelchiBelum ada peringkat
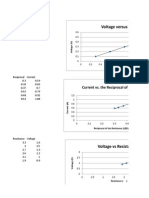
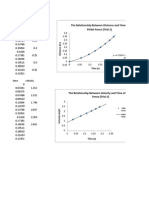
- The Relationship Between Distance and Time of Picket Fence (Trial 1)Dokumen2 halamanThe Relationship Between Distance and Time of Picket Fence (Trial 1)MelchiBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Chemical Kinetics TheoryDokumen31 halamanChemical Kinetics TheoryKivilia EduventuresBelum ada peringkat
- Chemistry PPT 21508Dokumen1 halamanChemistry PPT 21508Sunil SharmaBelum ada peringkat
- Water-Gas Shift Kinetics Over Iron Oxide Catalysts at Membrane Reactor ConditionsDokumen43 halamanWater-Gas Shift Kinetics Over Iron Oxide Catalysts at Membrane Reactor ConditionsshubhamBelum ada peringkat
- 1.3b Enzymes EditedDokumen10 halaman1.3b Enzymes EditedSoleil R.Belum ada peringkat
- NCERT Solutions For Class 12 Chemistry Chapter 4 Chemical KineticsDokumen31 halamanNCERT Solutions For Class 12 Chemistry Chapter 4 Chemical KineticsGourav ARBelum ada peringkat
- KineticsDokumen26 halamanKineticsMelissa M. Abansi-BautistaBelum ada peringkat
- Chem 1B Do Over EXAM 1Dokumen5 halamanChem 1B Do Over EXAM 1jarrodjohnBelum ada peringkat
- Chemical Kinetics: Aa BB CC DD + +Dokumen1 halamanChemical Kinetics: Aa BB CC DD + +Chandra Vamsi AdhikariBelum ada peringkat
- Study of The Apparent Kinetics of Biomass Gasification Using High-Temperature SteamDokumen55 halamanStudy of The Apparent Kinetics of Biomass Gasification Using High-Temperature SteamiqraSarfrazBelum ada peringkat
- Enzyme Kinetics Report FormatDokumen5 halamanEnzyme Kinetics Report FormatOnrcn BgntsBelum ada peringkat
- Sdarticle 004Dokumen85 halamanSdarticle 004geo angBelum ada peringkat
- Chem 444 Lab SyllabusDokumen100 halamanChem 444 Lab SyllabusStephano ParkBelum ada peringkat
- Worksheet 4 Moses Miller - Overview of Enzyme KineticsDokumen5 halamanWorksheet 4 Moses Miller - Overview of Enzyme Kineticsscience departmentBelum ada peringkat
- Chemical KineticsDokumen24 halamanChemical KineticsJason TyslBelum ada peringkat
- Kinetics of Gas-Liquid ReactionsDokumen7 halamanKinetics of Gas-Liquid ReactionsRajesh NimmakayalaBelum ada peringkat
- Polymath SimulationDokumen3 halamanPolymath SimulationToMemBelum ada peringkat
- Investigation - Ideas - Files - The Kinetics of The Iodination of Acetone PDFDokumen9 halamanInvestigation - Ideas - Files - The Kinetics of The Iodination of Acetone PDFkgothatso maleteBelum ada peringkat
- Ester HydrolysisDokumen3 halamanEster Hydrolysissheik abdullaBelum ada peringkat
- NitrationDokumen38 halamanNitrationHajra AamirBelum ada peringkat
- Grade-9 Chemistry WorksheetDokumen28 halamanGrade-9 Chemistry WorksheetKIdus Geta100% (1)
- Fluidized Bedreactor Modeling and SimulaDokumen13 halamanFluidized Bedreactor Modeling and Simulabader291Belum ada peringkat
- Nato - Stanag 4582 - Explosives, Nitrocellulose Based Propellants, Stability Test Procedure and Requirements Using Heat Flow CalorimetryDokumen25 halamanNato - Stanag 4582 - Explosives, Nitrocellulose Based Propellants, Stability Test Procedure and Requirements Using Heat Flow CalorimetryTetis BrionesBelum ada peringkat
- 1 s2.0 S0957582023002264 MainDokumen22 halaman1 s2.0 S0957582023002264 Mainlunar100% (1)
- Resfaculty 10Dokumen78 halamanResfaculty 10nandi_scrBelum ada peringkat
- ChemistryDokumen2 halamanChemistryGokul VishwaBelum ada peringkat
- 5.2 - Enzyme Parameters - Chemistry LibreTextsDokumen1 halaman5.2 - Enzyme Parameters - Chemistry LibreTextsteam TSOTAREBelum ada peringkat
- 0775.chemical Degradation Methods For Wastes and Pollutants. Environmental and Industrial Applications by Matthew A. Tarr PDFDokumen478 halaman0775.chemical Degradation Methods For Wastes and Pollutants. Environmental and Industrial Applications by Matthew A. Tarr PDFmichaelBelum ada peringkat
- 3 - MS - Chemical KineticsDokumen11 halaman3 - MS - Chemical KineticsJems ChaudharyBelum ada peringkat
- Pengukuran Berapa Pantas Tindak Balas Berlaku: Rate of ReactionDokumen8 halamanPengukuran Berapa Pantas Tindak Balas Berlaku: Rate of ReactionFazza RudyBelum ada peringkat
- Stability Constants of Metal Complexes: Section - CDokumen34 halamanStability Constants of Metal Complexes: Section - ChaumeoBelum ada peringkat