Anda mungkin juga menyukai
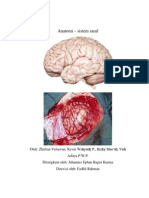
- Anatomi OtakDokumen18 halamanAnatomi OtakMuhammad Azhary LazuardyBelum ada peringkat
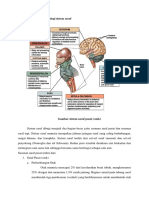
- Sistem SarafDokumen43 halamanSistem SarafJohannes CordeBelum ada peringkat
- Cerebrum .Dokumen18 halamanCerebrum .Afina Sabila100% (1)
- Anatomi Makroskopik Hippocampus Dan Hubungannya Dengan Struktur LainDokumen4 halamanAnatomi Makroskopik Hippocampus Dan Hubungannya Dengan Struktur LainCindy AmeLiyana Part IIBelum ada peringkat
- Telencephalon, Cerebellum, PonsDokumen37 halamanTelencephalon, Cerebellum, PonsHanda YaniBelum ada peringkat
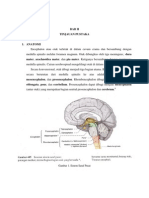
- Makalah Anatomi Sistem Saraf Pusat 1Dokumen19 halamanMakalah Anatomi Sistem Saraf Pusat 1Tiara Shifa AzzahraBelum ada peringkat
- Referat Sistem LimbikDokumen15 halamanReferat Sistem LimbikEdo Johanes SihombingBelum ada peringkat
- Struktur OtakDokumen10 halamanStruktur OtakFenita Renny DinataBelum ada peringkat
- Sistem NeurovaskulerDokumen21 halamanSistem NeurovaskulerVivitPutYapani0% (2)
- REFERATDokumen30 halamanREFERATIcha LightBelum ada peringkat
- Makalah ForensikDokumen37 halamanMakalah ForensikChrisnaArdhyaMedikaBelum ada peringkat
- OTAK DAN FUNGSINYADokumen8 halamanOTAK DAN FUNGSINYAbayuBelum ada peringkat
- Jurnal Pembelajaran 1.2 Tutorial HDokumen44 halamanJurnal Pembelajaran 1.2 Tutorial HKusnul AmaliaBelum ada peringkat
- Anatomi Sistem SarafDokumen7 halamanAnatomi Sistem SarafJane Irene JessicaBelum ada peringkat
- Traktus KortikospinalisDokumen8 halamanTraktus KortikospinalisFaisal RuhiatnaBelum ada peringkat
- Anatomi OtakDokumen11 halamanAnatomi OtakErik SantosoBelum ada peringkat
- EnsefalitisDokumen38 halamanEnsefalitisDenata PrabhasiwiBelum ada peringkat
- Handout Saraf 1234567Dokumen30 halamanHandout Saraf 1234567Narityastuti Dwi BudiBelum ada peringkat
- Stroke EmboliDokumen23 halamanStroke EmboliSri LindaBelum ada peringkat
- Bab II NadyaDokumen41 halamanBab II NadyaNadya Kuncaraning AnugraeBelum ada peringkat
- Lobus FrontalDokumen7 halamanLobus FrontalDexRizh Si Ceue AlayBelum ada peringkat
- 2003 Anatomi Hipotalamus Dan Kelenjar PituitariDokumen16 halaman2003 Anatomi Hipotalamus Dan Kelenjar PituitariNormanPrabowoBelum ada peringkat
- Fisiologi Saraf2Dokumen13 halamanFisiologi Saraf2Meilia DewiiBelum ada peringkat
- OTAKDokumen15 halamanOTAKkenangaBelum ada peringkat
- Traktus Kortikospinalis dan Fungsinya dalam Kontrol MotorikDokumen10 halamanTraktus Kortikospinalis dan Fungsinya dalam Kontrol MotorikAnonymous eLJcGbWBelum ada peringkat
- Truncus CerebriDokumen18 halamanTruncus CerebriSiFa Imah RhieshaBelum ada peringkat
- CER Et DienDokumen25 halamanCER Et DienBias HerkawentarBelum ada peringkat
- FISIOLOGIS OTAKDokumen5 halamanFISIOLOGIS OTAKImam AdliBelum ada peringkat
- Anatomi OtakDokumen3 halamanAnatomi Otakyessy simbolonBelum ada peringkat
- Nervujaras!Dokumen35 halamanNervujaras!Nuraga Wishnu PutraBelum ada peringkat
- Anatomi BLOK 21 PsikiatriDokumen18 halamanAnatomi BLOK 21 PsikiatriSyscha Lumempouw 'Kaine Agape'Belum ada peringkat
- Tugas Anatomi NeurologiDokumen83 halamanTugas Anatomi NeurologiMiss PramaysheillaBelum ada peringkat
- Anatomi Dan Fisiologi Nervus CranialisDokumen11 halamanAnatomi Dan Fisiologi Nervus CranialisaduhbingungBelum ada peringkat
- Referat - Ilham Dianugraha - 112018086Dokumen13 halamanReferat - Ilham Dianugraha - 112018086memmuse95Belum ada peringkat
- ANATOMIDokumen6 halamanANATOMITaufiq ZulyasmanBelum ada peringkat
- Anatomi Potong Lintang Kls 1bDokumen48 halamanAnatomi Potong Lintang Kls 1bchanyeolBelum ada peringkat
- Otak AnatomiDokumen8 halamanOtak Anatomi...Belum ada peringkat
- OtakDokumen75 halamanOtakIkhwan AsshiddqyBelum ada peringkat
- Anatomi Lobus ParietDokumen10 halamanAnatomi Lobus ParietRey naldi100% (1)
- Lobus Parietal dan OksipitalDokumen27 halamanLobus Parietal dan OksipitalArif rahmanBelum ada peringkat
- Skenario 4 Blok NeuroDokumen57 halamanSkenario 4 Blok NeuroNiken Audi LestariBelum ada peringkat
- Laporan Pendahuluan Sop Cerebri EdaDokumen25 halamanLaporan Pendahuluan Sop Cerebri EdaaprianiBelum ada peringkat
- Anatomi Dan Fisiologi Sistem LimbikDokumen13 halamanAnatomi Dan Fisiologi Sistem LimbikYoga Dananjaya67% (3)
- Sistem Saraf PusatDokumen66 halamanSistem Saraf Pusatpurb145Belum ada peringkat
- Kuliah Anatomi Blok 6. Cerebrum, Cerebellum Dan Sistem Limbik (Dr. Reni Oktavina, M.biomed) - DikonversiDokumen69 halamanKuliah Anatomi Blok 6. Cerebrum, Cerebellum Dan Sistem Limbik (Dr. Reni Oktavina, M.biomed) - DikonversiBeta BassistBelum ada peringkat
- Anatomi Dan FisiologiDokumen8 halamanAnatomi Dan Fisiologiintan_sari_25Belum ada peringkat
- Patofisiologi Modul KelumpuhanDokumen46 halamanPatofisiologi Modul KelumpuhanPirania Christy Tatipang-AndalangiBelum ada peringkat
- Struktur Otak BesarDokumen2 halamanStruktur Otak BesarApriliawatiBelum ada peringkat
- Anfis KepalaDokumen39 halamanAnfis KepalaMita AtmadjaBelum ada peringkat
- Referat - Sistem LimbikDokumen19 halamanReferat - Sistem Limbikm.m.m.mBelum ada peringkat
- SISTEM LIMBIKDokumen16 halamanSISTEM LIMBIKIda Bagus Dharma PutraBelum ada peringkat
- KMB STROKE Hemoragik BeresDokumen67 halamanKMB STROKE Hemoragik BeresZelika GzBelum ada peringkat
- Referat Lobus FrontalisDokumen18 halamanReferat Lobus FrontalisAnnisa Aprilia Athira100% (1)
- Refarat Traktus KortikospinalisDokumen15 halamanRefarat Traktus KortikospinalisAhmad Saiful100% (2)
- OtakDokumen12 halamanOtakkd dewiBelum ada peringkat
- Struktur Makroskopis Otak KiriDokumen3 halamanStruktur Makroskopis Otak KiriAnnisa NurfitriyaniBelum ada peringkat
- Kasus SAH Dalam Kehamilankehamilan111Dokumen42 halamanKasus SAH Dalam Kehamilankehamilan111Rafika MonalisaBelum ada peringkat
- Hub Status Demensia DG Disabilitas Funfsional LansiaDokumen12 halamanHub Status Demensia DG Disabilitas Funfsional LansiaRafika MonalisaBelum ada peringkat
- Abses SerebriDokumen21 halamanAbses SerebriRafika MonalisaBelum ada peringkat
- Case ICH in Pregnancy EditDokumen70 halamanCase ICH in Pregnancy EditRafika MonalisaBelum ada peringkat
- Translate Hub Sirkumsisi & ISKDokumen16 halamanTranslate Hub Sirkumsisi & ISKRafika MonalisaBelum ada peringkat
- PEMERIKSAAN NEUROLOGIDokumen55 halamanPEMERIKSAAN NEUROLOGImaria wahyuniBelum ada peringkat
- NEUROBEHAVIORDokumen58 halamanNEUROBEHAVIORRafika Monalisa100% (3)
- Syndroma ParaneoplastikDokumen31 halamanSyndroma ParaneoplastikRafika MonalisaBelum ada peringkat
- PPK (Panduan Praktek Klinik)Dokumen475 halamanPPK (Panduan Praktek Klinik)Ghiri Chocolatos100% (1)
- Isi Kasus Etik 1Dokumen24 halamanIsi Kasus Etik 1Patricia TalakuaBelum ada peringkat
- Modul - Neurovaskular PDFDokumen101 halamanModul - Neurovaskular PDFRafika MonalisaBelum ada peringkat
- TUGAS THT LOniDokumen26 halamanTUGAS THT LOnibraincheatsBelum ada peringkat
- HiperkoagulasipadaPasienKankerParuBukanSelKecil PDFDokumen6 halamanHiperkoagulasipadaPasienKankerParuBukanSelKecil PDFRafika MonalisaBelum ada peringkat
- Hubungan Senam Hamil Dengan Proses Dan Hasil Persalinan PrimiparaDokumen11 halamanHubungan Senam Hamil Dengan Proses Dan Hasil Persalinan PrimiparaRafika MonalisaBelum ada peringkat
- Proposal Ru III Akbar & Uda Revisi IaDokumen13 halamanProposal Ru III Akbar & Uda Revisi IaRafika MonalisaBelum ada peringkat
- Safe Motherhood Dan ANCDokumen15 halamanSafe Motherhood Dan ANCRafika MonalisaBelum ada peringkat
- Pemeriksaan IVA Dan Pap TesDokumen12 halamanPemeriksaan IVA Dan Pap TesDestri HandokoBelum ada peringkat
- KB Dan KontrasepsiDokumen15 halamanKB Dan KontrasepsiRafika MonalisaBelum ada peringkat
- Ukuran Frekuensi PenyakitDokumen23 halamanUkuran Frekuensi PenyakitRafika MonalisaBelum ada peringkat
- Mineral makro dan mikro untuk kesehatan tubuhDokumen27 halamanMineral makro dan mikro untuk kesehatan tubuhRafika MonalisaBelum ada peringkat
- StrokeRehabDokumen20 halamanStrokeRehabRafika MonalisaBelum ada peringkat
- Bab I PendahuluanDokumen31 halamanBab I PendahuluanRafika MonalisaBelum ada peringkat
- Portofolio Demam TifoidDokumen17 halamanPortofolio Demam TifoidRafika MonalisaBelum ada peringkat
- EMBRIOTOMIDokumen11 halamanEMBRIOTOMIRafika MonalisaBelum ada peringkat
- Kuretase Pasca PersalinanDokumen4 halamanKuretase Pasca PersalinanRafika MonalisaBelum ada peringkat