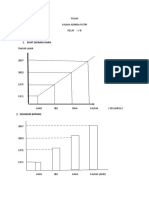
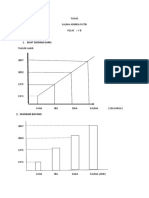
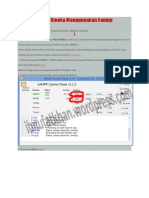
Anda mungkin juga menyukai
- Pengendalian Mikroorganisme Dengan Sarana Bahan KimiaDokumen30 halamanPengendalian Mikroorganisme Dengan Sarana Bahan Kimiarani noorBelum ada peringkat
- Kelompok 7 PteridophytaDokumen32 halamanKelompok 7 PteridophytaRara PororoBelum ada peringkat
- Basidiomycete SDokumen17 halamanBasidiomycete SRIyan Lare SaeBelum ada peringkat
- PCR (Polymerase Chain ReactionDokumen12 halamanPCR (Polymerase Chain ReactionLuthfi Hilman FBelum ada peringkat
- Karakteristik VirusDokumen7 halamanKarakteristik VirusAlit BioKunt100% (1)
- Alel Gnda AkuDokumen52 halamanAlel Gnda Akuputri pramitaBelum ada peringkat
- Plasmid, Episom dan Elemen Transposable BakteriDokumen5 halamanPlasmid, Episom dan Elemen Transposable Bakterifaris nizarBelum ada peringkat
- SPT - Sistem Simpodian Dan Anatomi NodalDokumen12 halamanSPT - Sistem Simpodian Dan Anatomi NodalAhmad RusdyBelum ada peringkat
- MACAM-MACAM MUTASI GENETIK DAN MUTASI YANG ACAKDokumen4 halamanMACAM-MACAM MUTASI GENETIK DAN MUTASI YANG ACAKFiqih Dewi Maharani100% (1)
- Preparat PollenDokumen5 halamanPreparat PollenardihaniBelum ada peringkat
- BAB 8rev-DikonversiDokumen31 halamanBAB 8rev-DikonversihomoriBelum ada peringkat
- MetamorfosisDokumen12 halamanMetamorfosisGovinda SitompulBelum ada peringkat
- Panduan Praktikum Botani Farmasi 2016-1Dokumen59 halamanPanduan Praktikum Botani Farmasi 2016-1sagitawidiyastuti0% (2)
- SPTDokumen7 halamanSPTAnonymous n8KNstNquBelum ada peringkat
- Klasifikasi Mikroba Dengan Taksonomi FILOGENETIKDokumen11 halamanKlasifikasi Mikroba Dengan Taksonomi FILOGENETIKPutri Intan MaharaniBelum ada peringkat
- Transkripsi Pada Prokariot ResumeDokumen3 halamanTranskripsi Pada Prokariot ResumeTiara Dwi NurmalitaBelum ada peringkat
- Evolusi KloroplasDokumen5 halamanEvolusi KloroplasAtika IndriyaniBelum ada peringkat
- Tumbuhan Dikotil Dan MonokotilDokumen3 halamanTumbuhan Dikotil Dan MonokotilDustin WilliamsBelum ada peringkat
- MorfogenesisDokumen25 halamanMorfogenesisIbu DaristasiBelum ada peringkat
- Laporan Ampertum 2 Uty FixDokumen14 halamanLaporan Ampertum 2 Uty FixPutri Mustika PratiwiBelum ada peringkat
- Agus Ulin Nuha 4411411011Dokumen5 halamanAgus Ulin Nuha 4411411011Agus Ulin NuhaBelum ada peringkat
- KONTROL ALAMIDokumen20 halamanKONTROL ALAMIria anitaBelum ada peringkat
- Pengelompokan BuahDokumen6 halamanPengelompokan BuahAldila RahmaBelum ada peringkat
- MAKALAH SPT Bunga KLP 3Dokumen19 halamanMAKALAH SPT Bunga KLP 3Rahma WatiBelum ada peringkat
- Rekayasa Genetika Untuk Produksi InterferonDokumen6 halamanRekayasa Genetika Untuk Produksi InterferonGhani Nurfiana100% (2)
- Apocynaceae Adalah Tumbuhan Berupa Pohon Atau SemakDokumen4 halamanApocynaceae Adalah Tumbuhan Berupa Pohon Atau SemakMifta NurzorifahBelum ada peringkat
- OOMYCETES: JAMUR AIR YANG SERING DIKELIRUKAN DENGAN FUNGIDokumen22 halamanOOMYCETES: JAMUR AIR YANG SERING DIKELIRUKAN DENGAN FUNGIrayhanBelum ada peringkat
- Laporan Praktikum 1 Mikrobiologi TerapanDokumen25 halamanLaporan Praktikum 1 Mikrobiologi Terapanahyar100% (2)
- SEL PROKARIOT DAN EUKARIOTDokumen10 halamanSEL PROKARIOT DAN EUKARIOTRivAy Orin SianipArBelum ada peringkat
- Kelompok 12 B (Makalah Virus Tumbuhan)Dokumen12 halamanKelompok 12 B (Makalah Virus Tumbuhan)fauzi cosmic100% (1)
- Ordo JuglandalesDokumen213 halamanOrdo JuglandalesUmmu Athiyyah0% (1)
- Morfologi Dan Struktur Sel Fungi 2Dokumen41 halamanMorfologi Dan Struktur Sel Fungi 2nurlatyfahfitrianingsihBelum ada peringkat
- DIAGRAM BUNGADokumen14 halamanDIAGRAM BUNGAgilangBelum ada peringkat
- Penentuan Jenis Kelamin Dan Rangkai KelaminDokumen3 halamanPenentuan Jenis Kelamin Dan Rangkai KelaminAnisa RahmalitaBelum ada peringkat
- Hipotesis Satu Gen Satu EnzimDokumen6 halamanHipotesis Satu Gen Satu EnzimAfif Saifudin100% (1)
- Materi NyaDokumen8 halamanMateri NyaIrma WahyuniastiBelum ada peringkat
- BRYOPHYTADokumen8 halamanBRYOPHYTAnurliazaharaBelum ada peringkat
- DNA Isolasi SederhanaDokumen7 halamanDNA Isolasi SederhanaSyafrizal UlumBelum ada peringkat
- Peran EcdysoneDokumen2 halamanPeran EcdysoneAidiel100% (1)
- 1-Evolusi Imunologi & KomparatifDokumen10 halaman1-Evolusi Imunologi & KomparatifFarradea Tiara PutriBelum ada peringkat
- REGULASI GEN PADA PERKEMBANGANDokumen8 halamanREGULASI GEN PADA PERKEMBANGANDhanial Fahira YasminBelum ada peringkat
- 7 Komp Non-ProtoplasmikDokumen21 halaman7 Komp Non-ProtoplasmikFitri Fitrotillah AbidinBelum ada peringkat
- Klasifikasi dan Morfologi Jasminum sambacDokumen10 halamanKlasifikasi dan Morfologi Jasminum sambacSusi LestariiBelum ada peringkat
- Vaksin RekombinanDokumen31 halamanVaksin RekombinanUrsula Herfina Tri KusumastutiBelum ada peringkat
- BAB IItugas Miko 1Dokumen25 halamanBAB IItugas Miko 1wulan dianiBelum ada peringkat
- Karakteristik, Reproduksi dan Peranan AscomycetesDokumen6 halamanKarakteristik, Reproduksi dan Peranan AscomycetesIddaa Daafiia OrchidaceaeBelum ada peringkat
- PERSILANGAN DIHIBRIDDokumen17 halamanPERSILANGAN DIHIBRIDUBijbaErgouna100% (2)
- SISTEM SELF INKOMPATIBILITAS TANAMANDokumen4 halamanSISTEM SELF INKOMPATIBILITAS TANAMANauliahasnaBelum ada peringkat
- Resume Mutasi Kromosom BAB 5Dokumen12 halamanResume Mutasi Kromosom BAB 5Rina Fiji LestariBelum ada peringkat
- Struktur dan Fungsi MitokondriaDokumen8 halamanStruktur dan Fungsi MitokondriaWiwienCaricaBelum ada peringkat
- Bryophyta Morsistum 1.1Dokumen13 halamanBryophyta Morsistum 1.1Fitri Andriani IbrahimBelum ada peringkat
- Klasifikasi Takson dan TingkatannyaDokumen3 halamanKlasifikasi Takson dan Tingkatannyadestri wijayantiBelum ada peringkat
- Laporan Praktikum Gerak BakteriDokumen21 halamanLaporan Praktikum Gerak BakteriIntan Sartika RiskyBelum ada peringkat
- Makalah Biologi MolekulerDokumen18 halamanMakalah Biologi MolekulerYunita ArasBelum ada peringkat
- Dinding Sel BakteriDokumen2 halamanDinding Sel BakteriYaniBelum ada peringkat
- Endosperm &embryo ReviewDokumen54 halamanEndosperm &embryo ReviewMuhammad FurqanBelum ada peringkat
- LHP 4 (Tasya Triana Putri - 1900017059)Dokumen3 halamanLHP 4 (Tasya Triana Putri - 1900017059)Tasya Triana PutriBelum ada peringkat
- Lac operon inducerDokumen10 halamanLac operon inducerRosi CwBelum ada peringkat
- Pengaturan Ekspresi Gen Pada ProkariotikDokumen10 halamanPengaturan Ekspresi Gen Pada ProkariotikismiatulBelum ada peringkat
- Regulasi Lac OperonDokumen14 halamanRegulasi Lac OperonM. SigitBelum ada peringkat
- Bahasa Indo 8Dokumen14 halamanBahasa Indo 8Danial Alfariez100% (1)
- Jawaban: B PembahasanDokumen22 halamanJawaban: B Pembahasanharyono iqbalBelum ada peringkat
- SiklusSelDokumen16 halamanSiklusSelharyono iqbalBelum ada peringkat
- Soal Tentang Sel UtamaDokumen4 halamanSoal Tentang Sel Utamaharyono iqbalBelum ada peringkat
- Di Bawah Ini Adalah KelainanDokumen8 halamanDi Bawah Ini Adalah Kelainanharyono iqbal100% (1)
- Tugas Diagram DindaDokumen2 halamanTugas Diagram Dindaharyono iqbalBelum ada peringkat
- PROPOSal Petaan Sekolah AsriDokumen13 halamanPROPOSal Petaan Sekolah Asriharyono iqbalBelum ada peringkat
- Bunga KemuningnyaDokumen2 halamanBunga Kemuningnyaharyono iqbalBelum ada peringkat
- Rencana Biologi SelDokumen43 halamanRencana Biologi Selharyono iqbalBelum ada peringkat
- Tugas Diagram DindaDokumen2 halamanTugas Diagram Dindaharyono iqbalBelum ada peringkat
- Proposal Green School ProgramDokumen13 halamanProposal Green School Programharyono iqbalBelum ada peringkat
- BDokumen13 halamanBharyono iqbalBelum ada peringkat
- Tutorial Install Omeka Menggunakan Xampp PadaDokumen6 halamanTutorial Install Omeka Menggunakan Xampp Padaharyono iqbalBelum ada peringkat
- Berita Acara Daftar HadirDokumen2 halamanBerita Acara Daftar Hadirharyono iqbalBelum ada peringkat
- MUTASIDokumen17 halamanMUTASIErlin WinarniiBelum ada peringkat
- 15Dokumen18 halaman15haryono iqbalBelum ada peringkat
- Pak Budi Menyilangkan Galur Murni Kacang Kapri Berbiji Bulat Warna KuningDokumen1 halamanPak Budi Menyilangkan Galur Murni Kacang Kapri Berbiji Bulat Warna Kuningharyono iqbal100% (1)
- Di Bawah Ini Adalah KelainanDokumen8 halamanDi Bawah Ini Adalah Kelainanharyono iqbal100% (1)
- Soal SPM UnDokumen9 halamanSoal SPM UnHaposan SiburianBelum ada peringkat
- Kumpulan Soal Tentang Reproduksi SelDokumen10 halamanKumpulan Soal Tentang Reproduksi Selharyono iqbalBelum ada peringkat
- BDokumen13 halamanBharyono iqbalBelum ada peringkat
- Pra Ujian Nasional Bahasa Inggris Sma Kode ADokumen17 halamanPra Ujian Nasional Bahasa Inggris Sma Kode Aharyono iqbalBelum ada peringkat
- 15Dokumen18 halaman15haryono iqbalBelum ada peringkat
- GENETIKADokumen19 halamanGENETIKAharyono iqbalBelum ada peringkat
- Soal Pewarisan SifatDokumen26 halamanSoal Pewarisan SifatGembiraLoka100% (1)
- Soal SPM UnDokumen9 halamanSoal SPM UnHaposan SiburianBelum ada peringkat
- JUDULDokumen13 halamanJUDULharyono iqbalBelum ada peringkat
- MATRIKSDokumen9 halamanMATRIKSNugraha SaputraBelum ada peringkat
- 13Dokumen14 halaman13haryono iqbalBelum ada peringkat