Anda mungkin juga menyukai
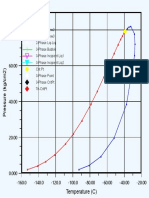
- Kurva SaprolDokumen1 halamanKurva SaprolTimotius Candra KusumaBelum ada peringkat
- Reynolds Average Navier-Stokes EquationDokumen108 halamanReynolds Average Navier-Stokes EquationTimotius Candra KusumaBelum ada peringkat
- Utility Systems: Course IntroductionDokumen4 halamanUtility Systems: Course IntroductionTimotius Candra KusumaBelum ada peringkat
- Gasal 1718 (FIX)Dokumen3 halamanGasal 1718 (FIX)Timotius Candra KusumaBelum ada peringkat
- Recombinant DnaDokumen23 halamanRecombinant DnaTimotius Candra KusumaBelum ada peringkat
- Accepted Manuscript: Journal of The Saudi Society of Agricultural SciencesDokumen15 halamanAccepted Manuscript: Journal of The Saudi Society of Agricultural SciencesTimotius Candra KusumaBelum ada peringkat
- Formulir Beasiswa YAGI - ITS PDFDokumen4 halamanFormulir Beasiswa YAGI - ITS PDFTimotius Candra KusumaBelum ada peringkat
- HiMedia Hugh Leifson Medium M826 Technical DatasheetDokumen2 halamanHiMedia Hugh Leifson Medium M826 Technical DatasheetTimotius Candra KusumaBelum ada peringkat
- Performance Test of Palm Fatty Acid Biodiesel On Compression Ignition EngineDokumen9 halamanPerformance Test of Palm Fatty Acid Biodiesel On Compression Ignition EngineTimotius Candra KusumaBelum ada peringkat
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Magic DrumDokumen185 halamanThe Magic Drumtanishgiri2012Belum ada peringkat
- ISO 9001 2015 AwarenessDokumen23 halamanISO 9001 2015 AwarenessSeni Oke0% (1)
- Farmer Producer Companies in OdishaDokumen34 halamanFarmer Producer Companies in OdishaSuraj GantayatBelum ada peringkat
- Ob NotesDokumen8 halamanOb NotesRahul RajputBelum ada peringkat
- of Thesis ProjectDokumen2 halamanof Thesis ProjectmoonBelum ada peringkat
- Test ScienceDokumen2 halamanTest Sciencejam syBelum ada peringkat
- Rule 113 114Dokumen7 halamanRule 113 114Shaila GonzalesBelum ada peringkat
- The Construction of Optimal Portfolio Using Sharpe's Single Index Model - An Empirical Study On Nifty Metal IndexDokumen9 halamanThe Construction of Optimal Portfolio Using Sharpe's Single Index Model - An Empirical Study On Nifty Metal IndexRevanKumarBattuBelum ada peringkat
- Parliament of India: Rajya SabhaDokumen64 halamanParliament of India: Rajya SabhaSivapothuraju KonathalaBelum ada peringkat
- DP 2 Human IngenuityDokumen8 halamanDP 2 Human Ingenuityamacodoudiouf02Belum ada peringkat
- Functional DesignDokumen17 halamanFunctional DesignRajivSharmaBelum ada peringkat
- History of The Sikhs by Major Henry Cour PDFDokumen338 halamanHistory of The Sikhs by Major Henry Cour PDFDr. Kamalroop SinghBelum ada peringkat
- Presentation 11Dokumen14 halamanPresentation 11stellabrown535Belum ada peringkat
- Problem Set-02Dokumen2 halamanProblem Set-02linn.pa.pa.khaing.2020.2021.fbBelum ada peringkat
- Sam Media Recruitment QuestionnaireDokumen17 halamanSam Media Recruitment Questionnairechek taiBelum ada peringkat
- Grade 7 ExamDokumen3 halamanGrade 7 ExamMikko GomezBelum ada peringkat
- Functions in C++Dokumen23 halamanFunctions in C++Abhishek ModiBelum ada peringkat
- Excon2019 ShowPreview02122019 PDFDokumen492 halamanExcon2019 ShowPreview02122019 PDFSanjay KherBelum ada peringkat
- Technical Specification For 33KV VCB BoardDokumen7 halamanTechnical Specification For 33KV VCB BoardDipankar ChatterjeeBelum ada peringkat
- Making Effective Powerpoint Presentations: October 2014Dokumen18 halamanMaking Effective Powerpoint Presentations: October 2014Mariam TchkoidzeBelum ada peringkat
- Obesity - The Health Time Bomb: ©LTPHN 2008Dokumen36 halamanObesity - The Health Time Bomb: ©LTPHN 2008EVA PUTRANTO100% (2)
- Aleksandrov I Dis 1-50.ru - enDokumen50 halamanAleksandrov I Dis 1-50.ru - enNabeel AdilBelum ada peringkat
- Turn Around Coordinator Job DescriptionDokumen2 halamanTurn Around Coordinator Job DescriptionMikeBelum ada peringkat
- E0 UoE Unit 7Dokumen16 halamanE0 UoE Unit 7Patrick GutierrezBelum ada peringkat
- Theory GraphDokumen23 halamanTheory GraphArthur CarabioBelum ada peringkat
- Skills Redux (10929123)Dokumen23 halamanSkills Redux (10929123)AndrewCollas100% (1)
- Chemistry: Crash Course For JEE Main 2020Dokumen18 halamanChemistry: Crash Course For JEE Main 2020Sanjeeb KumarBelum ada peringkat
- The Homework Song FunnyDokumen5 halamanThe Homework Song Funnyers57e8s100% (1)
- 74HC00D 74HC00D 74HC00D 74HC00D: CMOS Digital Integrated Circuits Silicon MonolithicDokumen8 halaman74HC00D 74HC00D 74HC00D 74HC00D: CMOS Digital Integrated Circuits Silicon MonolithicAssistec TecBelum ada peringkat
- 04 - Fetch Decode Execute Cycle PDFDokumen3 halaman04 - Fetch Decode Execute Cycle PDFShaun HaxaelBelum ada peringkat