Anda mungkin juga menyukai
- Sunfix Blue SPRDokumen7 halamanSunfix Blue SPRDyeing 2 Wintex100% (2)
- VIP45Dokumen92 halamanVIP45Román IsraelBelum ada peringkat
- Laboratory Experiment 3 Test For CarbohydratesDokumen9 halamanLaboratory Experiment 3 Test For CarbohydratesRenee Dwi Permata MessakaraengBelum ada peringkat
- The Aroids of Borneo.: January 2004Dokumen4 halamanThe Aroids of Borneo.: January 2004Negeri HijauBelum ada peringkat
- Cajaninae of Australia (Leguminosae: Papilionoideae) : L. J. G. Van Der MaesenDokumen9 halamanCajaninae of Australia (Leguminosae: Papilionoideae) : L. J. G. Van Der MaesenSheilla Moreno PérezBelum ada peringkat
- Colocasia Esculenta PDFDokumen20 halamanColocasia Esculenta PDFpaneisanBelum ada peringkat
- Spheciospongia Vesparium, Commonly Known As The LoggerheadDokumen2 halamanSpheciospongia Vesparium, Commonly Known As The Loggerheadandrew_yeap_2Belum ada peringkat
- Project 2019Dokumen24 halamanProject 2019tanmoy sardarBelum ada peringkat
- Basa3 PDFDokumen27 halamanBasa3 PDFJamal HuntBelum ada peringkat
- FoRAM Index Functional GroupsDokumen3 halamanFoRAM Index Functional GroupsGerard GomezBelum ada peringkat
- TheAroidsofBorneo Gardenwise231113 Boyce2004Dokumen4 halamanTheAroidsofBorneo Gardenwise231113 Boyce2004Faradlina MuftiBelum ada peringkat
- A Study On Species Diversity of Medicinal Plants in Ponmudi and Kallar LocalityDokumen13 halamanA Study On Species Diversity of Medicinal Plants in Ponmudi and Kallar LocalityanoojslBelum ada peringkat
- Vespula GermanicaDokumen7 halamanVespula GermanicaT NgBelum ada peringkat
- IN690Dokumen3 halamanIN690Sri MaulidiniBelum ada peringkat
- Caribbean AnolesDokumen68 halamanCaribbean AnolesNelly JiménezBelum ada peringkat
- AmaryllisDokumen4 halamanAmaryllistTEInjr0kZBelum ada peringkat
- Common, Tagalog, and Scientific Names of Fruits and VegetablesDokumen8 halamanCommon, Tagalog, and Scientific Names of Fruits and VegetablesNeahrabihagBelum ada peringkat
- Aristolochia FimbriataDokumen26 halamanAristolochia FimbriataDiego MyxomycotaBelum ada peringkat
- Amanita Muscaria - Wikipedia, The Free Encyclopedia PDFDokumen17 halamanAmanita Muscaria - Wikipedia, The Free Encyclopedia PDFAmeli ApuyBelum ada peringkat
- Aporometra Wilsoni: Aporometra Wilsoni Is A Marine Invertebrate, A Species of Crinoid orDokumen2 halamanAporometra Wilsoni: Aporometra Wilsoni Is A Marine Invertebrate, A Species of Crinoid orCarlos de la CallejaBelum ada peringkat
- Harris&02Dokumen5 halamanHarris&02sd24Belum ada peringkat
- Revision of The Genus Bromheadia (OrchidaceaeDokumen40 halamanRevision of The Genus Bromheadia (OrchidaceaeCherie Anne GrospeBelum ada peringkat
- Armillaria SolidipesDokumen9 halamanArmillaria SolidipesIzzah Akimoto AyumuBelum ada peringkat
- SCHlegeliaceae Nueva Especie para Del Bosque Montano Húmedo de Colombia y Una Clave para Las Especies Del GéneroDokumen13 halamanSCHlegeliaceae Nueva Especie para Del Bosque Montano Húmedo de Colombia y Una Clave para Las Especies Del GéneroFelipe KoyotlBelum ada peringkat
- Violet EarDokumen2 halamanViolet EarFelix100% (1)
- Rare Forest Plants Southern BCDokumen6 halamanRare Forest Plants Southern BCzineil BlackwoodBelum ada peringkat
- Species Orchidacearum Icones Colombianae 2 LCDP14 1Dokumen1 halamanSpecies Orchidacearum Icones Colombianae 2 LCDP14 1Sugey Ortiz-SerranoBelum ada peringkat
- 1 Honeybees in Nature-Thomas D. Seeley PDFDokumen11 halaman1 Honeybees in Nature-Thomas D. Seeley PDFDavid Ernesto Mayen MilianBelum ada peringkat
- LiliaceaeDokumen13 halamanLiliaceaesoumya vermaBelum ada peringkat
- The Origin and Domestication of Aquilaria, An Important Agarwood-Producing GenusDokumen20 halamanThe Origin and Domestication of Aquilaria, An Important Agarwood-Producing GenusLy KhánhBelum ada peringkat
- UnnamedDokumen35 halamanUnnamedSiva SivakumarBelum ada peringkat
- The Alstroemeriaceae in Peru and Neighbouring Areas: Anton Hofreiter and Eric F. RodríguezDokumen66 halamanThe Alstroemeriaceae in Peru and Neighbouring Areas: Anton Hofreiter and Eric F. RodríguezJonathan Urrutia EstradaBelum ada peringkat
- The Phylogeny of The Alstroemeriaceae, Based On Morphology, rps16 Intron, andDokumen24 halamanThe Phylogeny of The Alstroemeriaceae, Based On Morphology, rps16 Intron, andGabriela DíazBelum ada peringkat
- Anatomical Studies On The Genus Arabia (Subgenera:: Euphorbia L. Saudi Triucalli, Ermophyton, Esula and Chamaesyce)Dokumen24 halamanAnatomical Studies On The Genus Arabia (Subgenera:: Euphorbia L. Saudi Triucalli, Ermophyton, Esula and Chamaesyce)etab alriashiBelum ada peringkat
- Kanab Ambersnail: by Shane Johns NOVEMBER 21, 2008Dokumen1 halamanKanab Ambersnail: by Shane Johns NOVEMBER 21, 2008s.j120962028100% (2)
- STS BiodiversityDokumen5 halamanSTS BiodiversityAndrea OesmerBelum ada peringkat
- 4 2 38.1Dokumen6 halaman4 2 38.1Alonso Barrios TrillerasBelum ada peringkat
- Biological Control: A Novel Approach To Managing Himalayan BalsamDokumen3 halamanBiological Control: A Novel Approach To Managing Himalayan BalsamCABIBelum ada peringkat
- Species, Castes and Biology of Honey Bees: Amit ChoudharyDokumen41 halamanSpecies, Castes and Biology of Honey Bees: Amit Choudharyhappy singhBelum ada peringkat
- Berberis Aquifolium-PhytotherDokumen4 halamanBerberis Aquifolium-PhytotherLoyal DoerBelum ada peringkat
- BroadbillDokumen1 halamanBroadbillAnonymous F3jHpqBelum ada peringkat
- 101 Savill Wilson Mason Jinks Silver Firs QJFDokumen13 halaman101 Savill Wilson Mason Jinks Silver Firs QJFNVAzirBelum ada peringkat
- Banksia Oblongifolia: Banksia Oblongifolia, Commonly Known As The Fern-Leaved, Dwarf or RustyDokumen8 halamanBanksia Oblongifolia: Banksia Oblongifolia, Commonly Known As The Fern-Leaved, Dwarf or RustyLarisaOnigaBelum ada peringkat
- Bee Basics An Introduction To Our Native BeesDokumen51 halamanBee Basics An Introduction To Our Native BeesDivya ArunimaBelum ada peringkat
- Blue-Throated Barbet - Wikipedia, The Free EncyclopediaDokumen2 halamanBlue-Throated Barbet - Wikipedia, The Free Encyclopediaapi-20818983Belum ada peringkat
- American PaddlefishDokumen8 halamanAmerican PaddlefishMyfbook PageBelum ada peringkat
- Basic Taxonomic Concept-Module 2Dokumen2 halamanBasic Taxonomic Concept-Module 2gabrieeellqBelum ada peringkat
- Pollination Mechanisms and Plant-Pollinator Relationships: Taxonomy BasicsDokumen20 halamanPollination Mechanisms and Plant-Pollinator Relationships: Taxonomy Basicsmaxamadaxmad365Belum ada peringkat
- Ardipithecus RamidusDokumen5 halamanArdipithecus RamidusJavier Sanguino BracamonteBelum ada peringkat
- AmaranthusDokumen12 halamanAmaranthusDavidBelum ada peringkat
- Lily Flowering Plant Liliales: Liliaceae, TheDokumen12 halamanLily Flowering Plant Liliales: Liliaceae, Theabubakar shuwaBelum ada peringkat
- 3 - AroidsDokumen4 halaman3 - AroidsJuan Medina SanchezBelum ada peringkat
- Jenis-Jenis Araceae Di Hutan Wisata Dan Cagar Alam Plawangan - Turgo YogyakartaDokumen11 halamanJenis-Jenis Araceae Di Hutan Wisata Dan Cagar Alam Plawangan - Turgo YogyakartaFiniartiBelum ada peringkat
- Taxonomic Revision of Cinnamomum (Lauraceae) in Borneo: Soh Wuu-KuangDokumen24 halamanTaxonomic Revision of Cinnamomum (Lauraceae) in Borneo: Soh Wuu-KuangPatriciaVlBelum ada peringkat
- Taxonomic Revision of Cinnamomum (Lauraceae) in Borneo: Soh Wuu-KuangDokumen24 halamanTaxonomic Revision of Cinnamomum (Lauraceae) in Borneo: Soh Wuu-KuangPatriciaVlBelum ada peringkat
- Diversity ProjectDokumen2 halamanDiversity ProjectHimani Kondlahalli Channabasappa VeerendraBelum ada peringkat
- Https en - Wikipedia.org Wiki AlderDokumen13 halamanHttps en - Wikipedia.org Wiki AlderSebastian GhermanBelum ada peringkat
- 3 Meerow 1998Dokumen2 halaman3 Meerow 1998Emma ValentinaBelum ada peringkat
- Campylomma Verbasci (Meyer) : (Hemiptera: Miridae)Dokumen2 halamanCampylomma Verbasci (Meyer) : (Hemiptera: Miridae)farinoosh askariBelum ada peringkat
- Auricularia: The Wood Ear MushroomDokumen30 halamanAuricularia: The Wood Ear MushroomSusmita SettBelum ada peringkat
- 10 1600@036364419X697994Dokumen38 halaman10 1600@036364419X697994Juan David Rodriguez HurtadoBelum ada peringkat
- Todiras Cristian - 1 PDFDokumen1 halamanTodiras Cristian - 1 PDFConstantin TodiBelum ada peringkat
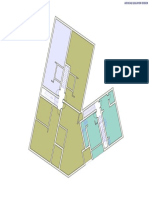
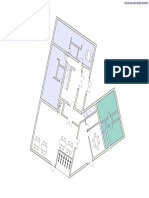
- Plan Mezanin Plan Etaj Curent Plan Etaj 6: Sali Administratie Cca. 88 MPDokumen1 halamanPlan Mezanin Plan Etaj Curent Plan Etaj 6: Sali Administratie Cca. 88 MPConstantin TodiBelum ada peringkat
- Plan de SituatieDokumen1 halamanPlan de SituatieConstantin TodiBelum ada peringkat
- 15R X 0.20 14G X 0.25: GseducationalversionDokumen1 halaman15R X 0.20 14G X 0.25: GseducationalversionConstantin TodiBelum ada peringkat
- 15R X 0.20 14G X 0.25: GseducationalversionDokumen1 halaman15R X 0.20 14G X 0.25: GseducationalversionConstantin TodiBelum ada peringkat
- Gseducationalversion: 15R X0. 20 14G X0. 25Dokumen1 halamanGseducationalversion: 15R X0. 20 14G X0. 25Constantin TodiBelum ada peringkat
- P+1 P+1 P+1 P+1: GseducationalversionDokumen1 halamanP+1 P+1 P+1 P+1: GseducationalversionConstantin TodiBelum ada peringkat
- Basement (Geology)Dokumen3 halamanBasement (Geology)Constantin TodiBelum ada peringkat
- P+1 P+1 P+1 P+1: GseducationalversionDokumen1 halamanP+1 P+1 P+1 P+1: GseducationalversionConstantin TodiBelum ada peringkat
- 11 Vernacular Building Techniques That Are DisappearingDokumen25 halaman11 Vernacular Building Techniques That Are DisappearingConstantin TodiBelum ada peringkat
- Gondwana (Dokumen14 halamanGondwana (Constantin TodiBelum ada peringkat
- S. Parcela: 585 MP P.O.T: 204 .8 MP C.U.T: 0.7 S. Parcela: 917 MP P.O.T: 321 MP C.U.T: 0.7Dokumen1 halamanS. Parcela: 585 MP P.O.T: 204 .8 MP C.U.T: 0.7 S. Parcela: 917 MP P.O.T: 321 MP C.U.T: 0.7Constantin TodiBelum ada peringkat
- BangladeshDokumen45 halamanBangladeshConstantin TodiBelum ada peringkat
- Synonym (Taxonomy)Dokumen5 halamanSynonym (Taxonomy)Constantin TodiBelum ada peringkat
- United States Patent (19) : U.S. Patent DocumentsDokumen6 halamanUnited States Patent (19) : U.S. Patent DocumentsConstantin TodiBelum ada peringkat
- Red Oaks Drive, Frankton, Queenstown-Lakes, Otago PDFDokumen1 halamanRed Oaks Drive, Frankton, Queenstown-Lakes, Otago PDFCintiah KenedyBelum ada peringkat
- Block-1 BLIS-03 Unit-2 PDFDokumen15 halamanBlock-1 BLIS-03 Unit-2 PDFravinderreddynBelum ada peringkat
- MalachiteDokumen2 halamanMalachiteAkhil KumarBelum ada peringkat
- MPI Report 009 Sub Structure B668Dokumen1 halamanMPI Report 009 Sub Structure B668John DolanBelum ada peringkat
- T60n02rg PDFDokumen8 halamanT60n02rg PDFsandor9116100% (2)
- Manual Elspec SPG 4420Dokumen303 halamanManual Elspec SPG 4420Bairon Alvira ManiosBelum ada peringkat
- Itinerary - State 2010Dokumen3 halamanItinerary - State 2010purest123Belum ada peringkat
- (Js-Umum) Daftar Harga 01 Maret '23Dokumen1 halaman(Js-Umum) Daftar Harga 01 Maret '23Kristin NataliaBelum ada peringkat
- NSTM.512v0.FANS.S9086 RS STM 010 CH 512R21Sep99Dokumen60 halamanNSTM.512v0.FANS.S9086 RS STM 010 CH 512R21Sep99jwfqcw74mfBelum ada peringkat
- 5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)Dokumen115 halaman5 - Oral Mucosa and Salivary Glands (Mahmoud Bakr)MobarobberBelum ada peringkat
- Traffic Cases in GSMDokumen18 halamanTraffic Cases in GSMkamal100% (1)
- RA - Ducting WorksDokumen6 halamanRA - Ducting WorksResearcherBelum ada peringkat
- UntitledDokumen300 halamanUntitledUnbox itBelum ada peringkat
- Athens 803 and The EkphoraDokumen18 halamanAthens 803 and The EkphoradovescryBelum ada peringkat
- Research On Strip Deformation in The Cage Roll-Forming Process of ERW Round PipesDokumen7 halamanResearch On Strip Deformation in The Cage Roll-Forming Process of ERW Round PipesJames PhillipsBelum ada peringkat
- NID DATPrelimsTestPaper2018 BDesDokumen24 halamanNID DATPrelimsTestPaper2018 BDesManaswini ReddyBelum ada peringkat
- 02K inDokumen1 halaman02K inAbbode HoraniBelum ada peringkat
- Indian Standard: Application Guide For Voltage TransformersDokumen16 halamanIndian Standard: Application Guide For Voltage TransformersGnanavel GBelum ada peringkat
- Backlash SDPDokumen11 halamanBacklash SDPPatrick Cyr GagnéBelum ada peringkat
- API 571 Quick ReviewDokumen32 halamanAPI 571 Quick ReviewMahmoud Hagag100% (1)
- Citadel of Kirkuk 1Dokumen17 halamanCitadel of Kirkuk 1ArézAzadBelum ada peringkat
- Regulatory Framework For Water Dams in QuebecDokumen2 halamanRegulatory Framework For Water Dams in QuebecRaveeBelum ada peringkat
- X Glo LED Strip Lighting For Tunnelling BrochureDokumen6 halamanX Glo LED Strip Lighting For Tunnelling BrochureJOSE HUAMANIBelum ada peringkat
- Soal Test All GrammarDokumen2 halamanSoal Test All GrammarAnonymous D7lnJMJYBelum ada peringkat
- Acc05 SCG116Dokumen42 halamanAcc05 SCG116Hilal HazaaBelum ada peringkat
- Procrustes AlgorithmDokumen11 halamanProcrustes AlgorithmShoukkathAliBelum ada peringkat
- The Practice of Veterinary Anesthesia - Small Animals, Birds, Fish, and ReptilesDokumen382 halamanThe Practice of Veterinary Anesthesia - Small Animals, Birds, Fish, and ReptilesMárcio MoreiraBelum ada peringkat