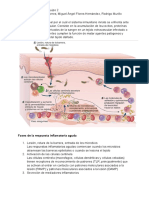
Anda mungkin juga menyukai
- Respuesta Adquirida (Inmunidad Adaptativa)Dokumen17 halamanRespuesta Adquirida (Inmunidad Adaptativa)Orbely Garcia100% (1)
- Funciones Del AmpcDokumen3 halamanFunciones Del AmpcRaulBelum ada peringkat
- Señalización Celular - Matriz ExtracelularDokumen7 halamanSeñalización Celular - Matriz ExtracelularEnzo Guillermo RamirezBelum ada peringkat
- RECEPTORESDokumen5 halamanRECEPTORESMaryen Gallardo ovandoBelum ada peringkat
- Compuestos NitrogenadosDokumen192 halamanCompuestos NitrogenadosMilton IsraelBelum ada peringkat
- Resumen-Fisiología de La Secreción PancreáticaDokumen3 halamanResumen-Fisiología de La Secreción PancreáticaJose Alfredo HernandezBelum ada peringkat
- La Enfermedad de Von GierkeDokumen10 halamanLa Enfermedad de Von GierkeKaren AtensioBelum ada peringkat
- Seminario de Celulas DendriticasDokumen24 halamanSeminario de Celulas DendriticasAnonymous IuimQGqnmBelum ada peringkat
- Resumen Guyton y Hall CAPITULO 81Dokumen11 halamanResumen Guyton y Hall CAPITULO 81Daniel Jaramillo AponteBelum ada peringkat
- MFH Iii - Ao 11Dokumen45 halamanMFH Iii - Ao 11CAngeles S Subero100% (1)
- Sindrome IctericoDokumen10 halamanSindrome IctericoDRIXBelum ada peringkat
- Agonistas ColinérgicosDokumen13 halamanAgonistas ColinérgicosruloBelum ada peringkat
- Inhibidor EnzimáticoDokumen18 halamanInhibidor EnzimáticoAdri CruzBelum ada peringkat
- Citoplasma y CitosolDokumen7 halamanCitoplasma y CitosolAlmacenCiberPreferidoBelum ada peringkat
- Resumen Inmunologia AcoDokumen135 halamanResumen Inmunologia AcoManuel González AlayesBelum ada peringkat
- Estructura Del Corpúsculo o Botón GustativoDokumen2 halamanEstructura Del Corpúsculo o Botón GustativoGustavo ArzolaBelum ada peringkat
- Fases de La GlucolisisDokumen7 halamanFases de La GlucolisisBraulio Castro'Belum ada peringkat
- Biosíntesis de Los AminoácidosDokumen4 halamanBiosíntesis de Los Aminoácidosmagda borjas100% (1)
- Sistema Renina - Angiotensina - Aldosterona 15Dokumen12 halamanSistema Renina - Angiotensina - Aldosterona 15R Martín Apresa MBelum ada peringkat
- Sistema Inmune 2018 Tipeo 2Dokumen34 halamanSistema Inmune 2018 Tipeo 2Belen Hernandez AguilarBelum ada peringkat
- Aparato de GolgiDokumen34 halamanAparato de GolgiMoisés J. Duran CanevaroBelum ada peringkat
- 02A Regulación MetabólicaDokumen40 halaman02A Regulación MetabólicaJhordy MamaniBelum ada peringkat
- Estructura CelularDokumen11 halamanEstructura CelularMaría José BarrenoBelum ada peringkat
- Desarrollo de Vellosidades CorionicasDokumen11 halamanDesarrollo de Vellosidades CorionicasAlexia AlvaradoBelum ada peringkat
- Metabolismo de Los CarbohidratosDokumen3 halamanMetabolismo de Los CarbohidratosMaria Melissa Tapias PerezBelum ada peringkat
- Proceso Inmunológico de La InflamaciónDokumen5 halamanProceso Inmunológico de La InflamaciónDavid Yepez100% (1)
- La Secreción de La InsulinaDokumen5 halamanLa Secreción de La Insulinaeloydc17Belum ada peringkat
- InflamaciónDokumen9 halamanInflamaciónIGNACIO SALVADOR FERNANDEZ GIRONBelum ada peringkat
- Resumen de Generalidades Del Sistema Nervioso.Dokumen7 halamanResumen de Generalidades Del Sistema Nervioso.Adriana ClarosBelum ada peringkat
- Hematopoyesis de Mecanismos de Ash RegulacionDokumen29 halamanHematopoyesis de Mecanismos de Ash Regulacionoliut74100% (1)
- Sintesis, Transporte y Excrecion de ColesterolDokumen4 halamanSintesis, Transporte y Excrecion de ColesterolPao Condo HidalgoBelum ada peringkat
- Cápsula InternaDokumen5 halamanCápsula InternaAnthony Romero100% (2)
- ArteroesclerosisDokumen72 halamanArteroesclerosisKatherine GallosoBelum ada peringkat
- Póster Aparato de GolgiDokumen1 halamanPóster Aparato de GolgiPaty RaazBelum ada peringkat
- EspermatogenesisDokumen12 halamanEspermatogenesisviriBelum ada peringkat
- Capítulo 15 - Señalización Celular y Transducción de Señales - Comunicación IntercelularDokumen75 halamanCapítulo 15 - Señalización Celular y Transducción de Señales - Comunicación IntercelularALEJANDRA INÉS NIETO ARIASBelum ada peringkat
- Enzimas Cardiacas CK CKMB Tgoasp LDHDokumen23 halamanEnzimas Cardiacas CK CKMB Tgoasp LDHAlberto GranadosBelum ada peringkat
- Eje Hipotalamo Hipofisis Ovario PresentacionDokumen25 halamanEje Hipotalamo Hipofisis Ovario PresentacionJesus CarreonBelum ada peringkat
- Exámenes Macroscópicos, Microscópicos y QuimicosDokumen4 halamanExámenes Macroscópicos, Microscópicos y QuimicosLuisaRodriguezBelum ada peringkat
- Transporte de ProteinasDokumen9 halamanTransporte de ProteinasGise MathierBelum ada peringkat
- División CelularDokumen10 halamanDivisión CelularAndrea Fernanda Calderon CuadrosBelum ada peringkat
- Semana 15 PNF MICDokumen11 halamanSemana 15 PNF MICKleimerContrerasBelum ada peringkat
- Asfixia Perinatal.Dokumen21 halamanAsfixia Perinatal.German Andres ZambranoBelum ada peringkat
- El EosinofiloDokumen4 halamanEl Eosinofilopepe islasBelum ada peringkat
- Seminario UnidoDokumen41 halamanSeminario UnidoAnonymous mv3B7mp1TYBelum ada peringkat
- CerebeloDokumen61 halamanCerebeloAlvaro Esteban FernándezBelum ada peringkat
- Sindrome de ZellwegerDokumen2 halamanSindrome de ZellwegerMaria Fabiola SantiagoBelum ada peringkat
- 8.1 Páncreas Endocrino PDFDokumen9 halaman8.1 Páncreas Endocrino PDFAna Cristina Ospina CairoBelum ada peringkat
- Capitulo 77 - Resumen Guyton e Hall - Fisiologia Medica 13 EdDokumen6 halamanCapitulo 77 - Resumen Guyton e Hall - Fisiologia Medica 13 Edgabriel valdiviaBelum ada peringkat
- Seminario #1 - Quimioprofilaxis y AntibióticosDokumen87 halamanSeminario #1 - Quimioprofilaxis y AntibióticosDamaris Guerra CarbonellBelum ada peringkat
- Bioqumica2 140307082934 Phpapp01Dokumen21 halamanBioqumica2 140307082934 Phpapp01Julimar FloresBelum ada peringkat
- Sintesis de Proteinas IVDokumen33 halamanSintesis de Proteinas IVLesly ResendizBelum ada peringkat
- Fagocitosis (Microbiología)Dokumen3 halamanFagocitosis (Microbiología)Andrea Huerta EscobedoBelum ada peringkat
- Cuestionario de BioquimicaDokumen8 halamanCuestionario de BioquimicaBel OcBelum ada peringkat
- FÍSIOLOGÍADokumen67 halamanFÍSIOLOGÍADaniela HinojosaBelum ada peringkat
- SeñalizacionDokumen4 halamanSeñalizacionFranciscoBelum ada peringkat
- Fisiologia Semana 4Dokumen11 halamanFisiologia Semana 4nicollgarsomBelum ada peringkat
- Com Intercelular 2Dokumen46 halamanCom Intercelular 2Natália LarsenBelum ada peringkat
- Segundos MensajerosDokumen42 halamanSegundos MensajerosMitch Peraza80% (10)
- Recepción Celular. Mecanismos MolecularesDokumen17 halamanRecepción Celular. Mecanismos MolecularesTEFABelum ada peringkat
- MM 0802020100Dokumen6 halamanMM 0802020100Luis Rafael CesarBelum ada peringkat
- LumbagoDokumen12 halamanLumbagoMacarena Vidal Illanes100% (1)
- Evaluacion Trabajo en CalienteDokumen2 halamanEvaluacion Trabajo en Calientegonzors0% (1)
- PLano Flujos A ColorDokumen1 halamanPLano Flujos A ColorJeremy Dolores MéndezBelum ada peringkat
- Informe 2. Sistema Masa-Resorte 1Dokumen4 halamanInforme 2. Sistema Masa-Resorte 1Leandra ÁlvarezBelum ada peringkat
- Historia de La Tabla PeriódicaDokumen9 halamanHistoria de La Tabla PeriódicaGuido Garnica ParedesBelum ada peringkat
- CarolinaDokumen18 halamanCarolinaCristofer Miñan EspinozaBelum ada peringkat
- S1.2 Actividad Aplicativa 02 Caso Shell - ESPDokumen7 halamanS1.2 Actividad Aplicativa 02 Caso Shell - ESPVladimir Noriega ChingBelum ada peringkat
- Antecedentes de La Norma Iso 22000Dokumen64 halamanAntecedentes de La Norma Iso 22000Jhonny Gonsalez100% (1)
- 0ejercicios 8 y 9 de Teoria Economica-PatatabravaDokumen12 halaman0ejercicios 8 y 9 de Teoria Economica-PatatabravaPaola HERNANDEZ GOMEZBelum ada peringkat
- Primeros Auxilios 2Dokumen12 halamanPrimeros Auxilios 2HBZ385Belum ada peringkat
- Otras Variables de MedicionDokumen18 halamanOtras Variables de MedicionRICARDO ROMEROBelum ada peringkat
- Proyecto Del Martillo MultiusosDokumen23 halamanProyecto Del Martillo Multiusosalejandra_sanchez_meza0% (2)
- Articulo La Presion Diferencial en Las Salas BlancasDokumen2 halamanArticulo La Presion Diferencial en Las Salas BlancasHector Hernan GimenezBelum ada peringkat
- Ética y ValoresDokumen2 halamanÉtica y ValoresMauricio Alejandro Arriagada HerazoBelum ada peringkat
- Trabajo Grupal - Semana 6Dokumen6 halamanTrabajo Grupal - Semana 6fernando jose carbajal carbajalBelum ada peringkat
- DESENTERRANDO A LOS ANCESTROS: LOS ORÍGENES DE LA PALEÓNTOLOGÍA HUMANA - Francisco Pelayo. Instituto López Piñero. CSIC. ValenciaDokumen22 halamanDESENTERRANDO A LOS ANCESTROS: LOS ORÍGENES DE LA PALEÓNTOLOGÍA HUMANA - Francisco Pelayo. Instituto López Piñero. CSIC. ValenciaFundación Canaria Orotava de Historia de la CienciaBelum ada peringkat
- Emisiones de Las Formas 1Dokumen4 halamanEmisiones de Las Formas 1GabrielInsua100% (2)
- Reducción de Una Carga Simple DistribuidaDokumen2 halamanReducción de Una Carga Simple DistribuidaJOSE ADRIAN MONTENEGRO QUESADABelum ada peringkat
- Falla Tipica de Nissan - Urban (Acelerado) (Solucion)Dokumen30 halamanFalla Tipica de Nissan - Urban (Acelerado) (Solucion)eiochoaBelum ada peringkat
- Bitacoras de Temperaturas Refrigerador EME 2Dokumen2 halamanBitacoras de Temperaturas Refrigerador EME 2OCTAVIO JARDINEZBelum ada peringkat
- ETICADokumen10 halamanETICARoger MooBelum ada peringkat
- Folleto Hyundai HD270 Volquete MixerDokumen2 halamanFolleto Hyundai HD270 Volquete MixerFredis Ospino altamar100% (1)
- Practica 1 - TrigoDokumen1 halamanPractica 1 - TrigoMARIO TEODOSIO ASTOS LAROTABelum ada peringkat
- Plan de Mejora de Los Aprendizajes de Tercer Grado 2021-81700Dokumen17 halamanPlan de Mejora de Los Aprendizajes de Tercer Grado 2021-81700Victoria Gonzalez Martell100% (1)
- Guia 1 Funcion de Nutrición y Cadenas y Redes TróficasDokumen6 halamanGuia 1 Funcion de Nutrición y Cadenas y Redes Tróficassebastian ochoaBelum ada peringkat
- Escogiendo Lo Mejor PDFDokumen63 halamanEscogiendo Lo Mejor PDFDicin G GuerraBelum ada peringkat
- RoboTech Storming Into The U.S. MarketDokumen12 halamanRoboTech Storming Into The U.S. MarketA_espinoBelum ada peringkat
- Examen de Los OjosDokumen9 halamanExamen de Los OjosCatalina VillanuevaBelum ada peringkat
- Ficha Evaluacion Entre Pares 2022Dokumen2 halamanFicha Evaluacion Entre Pares 2022Emerson Wilgen Apaza LopezBelum ada peringkat