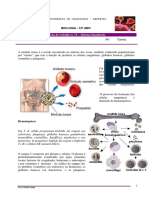
Anda mungkin juga menyukai
- Integração Do Metabolismo IntermediárioDokumen27 halamanIntegração Do Metabolismo IntermediárioMatheus Borges100% (1)
- GLICONEOGÊNESEDokumen10 halamanGLICONEOGÊNESESabrina SousaBelum ada peringkat
- Glícolise e gliconeogêneseDokumen8 halamanGlícolise e gliconeogêneseCharles Lameira ValenteBelum ada peringkat
- Glicólise - BQM IIDokumen5 halamanGlicólise - BQM IIJAQUELINE AGUIARBelum ada peringkat
- Bioquímica II - SimuladoDokumen7 halamanBioquímica II - SimuladoManu DenckBelum ada peringkat
- Metabolismo EnergéticoDokumen7 halamanMetabolismo EnergéticoMariana PaesBelum ada peringkat
- GlicogêneseDokumen5 halamanGlicogêneseLuanaBelum ada peringkat
- Resumos de BioquimicaDokumen42 halamanResumos de BioquimicaAna Carolina Soares EstevesBelum ada peringkat
- Gliconeogenese e Metabolismo de CarboidratosDokumen12 halamanGliconeogenese e Metabolismo de CarboidratosSuellenKartalian100% (1)
- Metabolismo dos Carboidratos: Digestão e Vias MetabólicasDokumen4 halamanMetabolismo dos Carboidratos: Digestão e Vias MetabólicasBia NunesBelum ada peringkat
- Gliconeogenese e Metabolismo Do GlicogênioDokumen27 halamanGliconeogenese e Metabolismo Do GlicogênioRicardo BorgesBelum ada peringkat
- Aula 22 - Neoglicogênese, Ciclo de Cori e de AlaninaDokumen22 halamanAula 22 - Neoglicogênese, Ciclo de Cori e de AlaninaAndré Jason MendesBelum ada peringkat
- Atividade GliconeogêneseDokumen5 halamanAtividade GliconeogêneseGatodo FabaBelum ada peringkat
- A GlicóliseDokumen4 halamanA GlicóliseAna Paula AzevedoBelum ada peringkat
- Processo de lipogênese e biossíntese de ácidos graxosDokumen5 halamanProcesso de lipogênese e biossíntese de ácidos graxosKAROLINE FAVORETO100% (1)
- A GlicóliseDokumen4 halamanA GlicóliseAna Paula AzevedoBelum ada peringkat
- Biossintese de Carboidratos BioquímicaDokumen6 halamanBiossintese de Carboidratos BioquímicaMarcela Lima de BritoBelum ada peringkat
- Metabolismo de LipídiosDokumen16 halamanMetabolismo de LipídiosDanielle RodriguesBelum ada peringkat
- Metabolismo Do GlicogênioDokumen6 halamanMetabolismo Do GlicogênioSara EsterBelum ada peringkat
- Lista de Exercicios Aula 13-Ciclo de Krebs e GlicogênioDokumen7 halamanLista de Exercicios Aula 13-Ciclo de Krebs e GlicogênioMaria MoreiraBelum ada peringkat
- A Gliconeogênese: Síntese de Glicose a Partir de PiruvatoDokumen21 halamanA Gliconeogênese: Síntese de Glicose a Partir de PiruvatoEveny PereiraBelum ada peringkat
- Metabolismo energético: piruvato e glicóliseDokumen10 halamanMetabolismo energético: piruvato e glicóliseVanessa Meirelles100% (2)
- Aula 14 - GluconeogéneseDokumen24 halamanAula 14 - GluconeogéneseaptgaptgBelum ada peringkat
- RevisãoDokumen6 halamanRevisãoe236610Belum ada peringkat
- Tema 2 - Glicólise, Gliconeogênese, e A Via Das Pentoses-FosfatoDokumen17 halamanTema 2 - Glicólise, Gliconeogênese, e A Via Das Pentoses-FosfatoDaniel FreireBelum ada peringkat
- Metabolismo de CarboidratosDokumen8 halamanMetabolismo de CarboidratosGabriel FerreiraBelum ada peringkat
- Tarefa de BioquimicaDokumen6 halamanTarefa de BioquimicaLucas CortezeBelum ada peringkat
- Neoglicogênese: síntese de glicose no fígado e rinsDokumen4 halamanNeoglicogênese: síntese de glicose no fígado e rinsMary FreitasBelum ada peringkat
- Microsoft Word - Estudo Dirigido - Bloco LipídiosDokumen4 halamanMicrosoft Word - Estudo Dirigido - Bloco LipídiosFelipe MagalhaesBelum ada peringkat
- Bioquimica MetabólicaDokumen4 halamanBioquimica MetabólicaLilian BertoluciBelum ada peringkat
- Bioquímica Síntese Da GlicoseDokumen9 halamanBioquímica Síntese Da GlicoseRê SilvaBelum ada peringkat
- Gliconeogenese e Via Das Pentoses FosfatosDokumen33 halamanGliconeogenese e Via Das Pentoses Fosfatoscarolaine santos100% (1)
- Estudo Dirigido BioquimicaDokumen5 halamanEstudo Dirigido Bioquimicaadelsonribeiro100% (4)
- Uma Panorâmica Geral Das Vias Metabólicas AvançadissimoDokumen4 halamanUma Panorâmica Geral Das Vias Metabólicas AvançadissimoallanfradiqueBelum ada peringkat
- Metabolismo de lipídeos: β-oxidação, corpos cetônicos e sínteseDokumen20 halamanMetabolismo de lipídeos: β-oxidação, corpos cetônicos e sínteseVanderval SIlva de OliveiraBelum ada peringkat
- Gliconeogênese: formação de glicose a partir de não-carboidratosDokumen25 halamanGliconeogênese: formação de glicose a partir de não-carboidratosAnderson SilveiraBelum ada peringkat
- Bioquimica Prova 1 PDFDokumen10 halamanBioquimica Prova 1 PDFESTHER DE MELLO ANTONIO100% (1)
- Uma Panorâmica Geral Das Vias MetabólicasDokumen4 halamanUma Panorâmica Geral Das Vias MetabólicasVanessa TomazBelum ada peringkat
- Metabolismo de carboidratos: gliconeogênese e regulação hormonalDokumen20 halamanMetabolismo de carboidratos: gliconeogênese e regulação hormonalAna SilvaBelum ada peringkat
- Glicogênese, Glicenolise, GliconeogeneseDokumen3 halamanGlicogênese, Glicenolise, GliconeogeneseLaryssa KlugeBelum ada peringkat
- Revisão p2 de BoinhaDokumen45 halamanRevisão p2 de BoinhaGlenda FrançaBelum ada peringkat
- Gluconeogênese: fonte glicose jejumDokumen3 halamanGluconeogênese: fonte glicose jejumHermes Sales CamposBelum ada peringkat
- GliconeogeneseDokumen4 halamanGliconeogeneseAline PereiraBelum ada peringkat
- Tutoria VitaminasDokumen18 halamanTutoria Vitaminas002-025384Belum ada peringkat
- Ciclo de Krebs e Glioxilato: metabolismo energético e alternativoDokumen2 halamanCiclo de Krebs e Glioxilato: metabolismo energético e alternativoTiemy Letícia ZanetteBelum ada peringkat
- Bioquímica Fisiológica Exame B - 210129 - 103356Dokumen5 halamanBioquímica Fisiológica Exame B - 210129 - 103356Joana Carolina Teixeira RibeiroBelum ada peringkat
- Bioquimica Metabolismo Integrado 09n 2Dokumen21 halamanBioquimica Metabolismo Integrado 09n 2Luciene Luba100% (2)
- Relatório 2 - Bioquímica 2 - Dosagem de Glicose e Fermentação em Saccharomyces CerevisiaeDokumen22 halamanRelatório 2 - Bioquímica 2 - Dosagem de Glicose e Fermentação em Saccharomyces CerevisiaeJheyce Moraes0% (1)
- BioquímicaDokumen5 halamanBioquímicaf252622Belum ada peringkat
- Bioquímica - Catabolismo de AminoácidosDokumen7 halamanBioquímica - Catabolismo de AminoácidosMarcela Lima de BritoBelum ada peringkat
- Mitocôndria desacoplada e termogêneseDokumen8 halamanMitocôndria desacoplada e termogêneseTatiane MoraisBelum ada peringkat
- Bioquimica - MetabolismoDokumen21 halamanBioquimica - MetabolismoNelma Assis Siqueira100% (1)
- IntegraçãoDokumen6 halamanIntegraçãoErika BennaiaBelum ada peringkat
- Resumos BioquímicaDokumen14 halamanResumos BioquímicaGabrieli HoepersBelum ada peringkat
- Carboidratos: conceitos, digestão, absorção e metabolismoDokumen20 halamanCarboidratos: conceitos, digestão, absorção e metabolismoAline da Silva Fernandes MatiasBelum ada peringkat
- Carboidratos - MetabolismoDokumen108 halamanCarboidratos - MetabolismoMathews Oviedo100% (1)
- Carboidratos, obesidade, glicogênio e metabolismo energéticoDokumen10 halamanCarboidratos, obesidade, glicogênio e metabolismo energéticoLucas KallielBelum ada peringkat
- Prolactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoDari EverandProlactina e Diabetes Melito do tipo 2: o efeito protetor de um hormônio sobre o metabolismo glicídicoBelum ada peringkat
- Cap 11 - Metabolismo Do GlicogênioDokumen3 halamanCap 11 - Metabolismo Do Glicogêniojbod2012100% (1)
- Fibroblastos e tipos de fibras do tecido conjuntivoDokumen2 halamanFibroblastos e tipos de fibras do tecido conjuntivojbod2012100% (1)
- Genética básica: revisão de conceitos fundamentaisDokumen16 halamanGenética básica: revisão de conceitos fundamentaisjbod2012Belum ada peringkat
- Resumo GeneticaDokumen10 halamanResumo Geneticajbod2012Belum ada peringkat
- Carburadores Vicente, MCDokumen43 halamanCarburadores Vicente, MCjbod2012Belum ada peringkat
- Samsung WB850FDokumen187 halamanSamsung WB850Fjbod2012Belum ada peringkat
- Melhorias Carburador 2e 3e menosDokumen5 halamanMelhorias Carburador 2e 3e menosjbod2012Belum ada peringkat
- Manualdoproesp BR 2012Dokumen20 halamanManualdoproesp BR 2012Nicole AiraghiBelum ada peringkat
- 4E-Ciclo de CoriDokumen6 halaman4E-Ciclo de Coriapi-2744148193% (15)
- Aula Prática de ImunologiaDokumen3 halamanAula Prática de ImunologiaLuiza LimaBelum ada peringkat
- Ebook Fisiologia HipertrofiaDokumen89 halamanEbook Fisiologia HipertrofiadstvguilandeBelum ada peringkat
- Aula 3.9 Beta - Oxidação RegulaçãoDokumen17 halamanAula 3.9 Beta - Oxidação Regulaçãowalas joãoBelum ada peringkat
- Tipos AasDokumen5 halamanTipos AasRafael RegatBelum ada peringkat
- Lista de revisão de imunologiaDokumen8 halamanLista de revisão de imunologialucas pazesBelum ada peringkat
- 11284816022012bioquimica Aula 13 PDFDokumen18 halaman11284816022012bioquimica Aula 13 PDFDiego PereiraBelum ada peringkat
- Integração metabólica durante jejum e pós-prandialDokumen34 halamanIntegração metabólica durante jejum e pós-prandialVinicius SoaresBelum ada peringkat
- Glicólise e ciclo de KrebsDokumen28 halamanGlicólise e ciclo de KrebsAvelino CardozoBelum ada peringkat
- Sistema Imunitário SecundárioDokumen5 halamanSistema Imunitário SecundárioMaria Maia100% (1)
- The Cube W1Dokumen11 halamanThe Cube W1Arthurdrummer1Belum ada peringkat
- Anatomia Do Sistema LinfáticoDokumen25 halamanAnatomia Do Sistema Linfáticopaulo67% (3)
- Questões metabólicas glicídicasDokumen5 halamanQuestões metabólicas glicídicasSergio AraujoBelum ada peringkat
- Células e Tecidos Do Sistema ImuneDokumen29 halamanCélulas e Tecidos Do Sistema ImunejosueeumesmoBelum ada peringkat
- PGCAN-HEMOG 06489 Flocky 421 MichelDokumen1 halamanPGCAN-HEMOG 06489 Flocky 421 MichelMichel PetersonBelum ada peringkat
- Resultado - 2023-06-28T231651.377Dokumen5 halamanResultado - 2023-06-28T231651.377daiane beneditoBelum ada peringkat
- Exames laboratoriais: taxas normais de creatinina, potássio, TGO, TGP e maisDokumen11 halamanExames laboratoriais: taxas normais de creatinina, potássio, TGO, TGP e maisOsias De Barros AnunciaçãoBelum ada peringkat
- Aula Células e Órgãos LinfoidesDokumen138 halamanAula Células e Órgãos LinfoidesNatan Gustavo100% (1)
- Exames laboratoriais de Cleber da Silva TellesDokumen6 halamanExames laboratoriais de Cleber da Silva TellesCarlos Daniel da Silva AlonsoBelum ada peringkat
- Metabolismo de Ácidos Graxos EstudoDokumen3 halamanMetabolismo de Ácidos Graxos EstudoRaquel Ramos Morita 202010257811100% (1)
- Ciclo Krebs SlidesDokumen31 halamanCiclo Krebs SlidesJoao Vitor de Oliveira FelixBelum ada peringkat
- Ciclo Dos Acidos Tricarboxilico - NutDokumen20 halamanCiclo Dos Acidos Tricarboxilico - Nutyasmim abreuBelum ada peringkat
- Bioquímica Aplicada À Saúde-Unidade 2-2.2-1ºsem2022-FolhetosDokumen15 halamanBioquímica Aplicada À Saúde-Unidade 2-2.2-1ºsem2022-FolhetosKauanna MartinsBelum ada peringkat
- Bioenergética e metabolismoDokumen22 halamanBioenergética e metabolismoBernardo Rafael Bittencourt BernardiBelum ada peringkat
- Aula 22 - Neoglicogênese, Ciclo de Cori e de AlaninaDokumen22 halamanAula 22 - Neoglicogênese, Ciclo de Cori e de AlaninaAndré Jason MendesBelum ada peringkat
- Resumo Obtenção de Energia Pelos Seres HeterotróficosDokumen15 halamanResumo Obtenção de Energia Pelos Seres HeterotróficosAna Beatriz AlmeidaBelum ada peringkat
- Sistema linfático: funções e circulação da linfaDokumen50 halamanSistema linfático: funções e circulação da linfaLeticiaBelum ada peringkat
- 19 Métodos e Sistemas de Treinamento de Força e Hipertrofia Muscular PDFDokumen8 halaman19 Métodos e Sistemas de Treinamento de Força e Hipertrofia Muscular PDFTimidoBelum ada peringkat
- Bioquímica Ii - Fermentação AlcoólicaDokumen149 halamanBioquímica Ii - Fermentação AlcoólicaMateus Metler SilvaBelum ada peringkat
- Metabolismo do glicogênio: síntese, degradação e regulaçãoDokumen49 halamanMetabolismo do glicogênio: síntese, degradação e regulaçãowanubio0% (1)