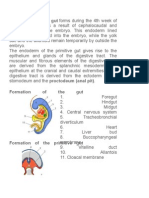
Anda mungkin juga menyukai
- Interstitial Collagen Synthesis and Processing in Human Amnion: A Property of The Mesenchymal Cells'Dokumen8 halamanInterstitial Collagen Synthesis and Processing in Human Amnion: A Property of The Mesenchymal Cells'Hòa HồBelum ada peringkat
- Infant Skin Barrier, Structure, and Enzymatic Activity Differ From Those of Adult in An East Asian CohortDokumen8 halamanInfant Skin Barrier, Structure, and Enzymatic Activity Differ From Those of Adult in An East Asian CohortS FznsBelum ada peringkat
- Stem Cell Therapy On Skin: Mechanisms, Recent Advances and Drug Reviewing IssuesDokumen7 halamanStem Cell Therapy On Skin: Mechanisms, Recent Advances and Drug Reviewing IssuesCandra BumiBelum ada peringkat
- Vanderputte 2011Dokumen8 halamanVanderputte 2011adamBelum ada peringkat
- 252 FullDokumen11 halaman252 FullshrutiBelum ada peringkat
- 1 s2.0 S0022202X15447736 MainDokumen7 halaman1 s2.0 S0022202X15447736 MainKhan FatemaBelum ada peringkat
- Mouse Epidermal Development - Effects of Retinoic Acid Exposure in Utero (Pages 36-44)Dokumen9 halamanMouse Epidermal Development - Effects of Retinoic Acid Exposure in Utero (Pages 36-44)jenBelum ada peringkat
- Articol DecelularizareDokumen12 halamanArticol DecelularizareOana MariaBelum ada peringkat
- Review Article: A Review of Fetal Scarless HealingDokumen10 halamanReview Article: A Review of Fetal Scarless Healingqaz1122Belum ada peringkat
- Articulo PaparpadoDokumen8 halamanArticulo PaparpadoJONATHAN CEDEÑOBelum ada peringkat
- Ahmad 2007Dokumen11 halamanAhmad 2007Erick MartinezBelum ada peringkat
- Comparison and Culturing Different Types of Cells From Fresh Breast Milk With Different Culture MediumDokumen9 halamanComparison and Culturing Different Types of Cells From Fresh Breast Milk With Different Culture Mediummutiara firdausyBelum ada peringkat
- Jean M. Lawrence Et Al - MIO-M1 Cells and Similar Muller Glial Cell Lines Derived From Adult Human Retina Exhibit Neural Stem Cell CharacteristicsDokumen12 halamanJean M. Lawrence Et Al - MIO-M1 Cells and Similar Muller Glial Cell Lines Derived From Adult Human Retina Exhibit Neural Stem Cell CharacteristicsHutsDMBelum ada peringkat
- Corneal Epithelial Proliferation and Thickness in A Mouse Model of Dry EyeDokumen7 halamanCorneal Epithelial Proliferation and Thickness in A Mouse Model of Dry EyeAa 1989Belum ada peringkat
- 1970 - Kollar - DP Embrionic FormationDokumen5 halaman1970 - Kollar - DP Embrionic FormationalexantonjazzBelum ada peringkat
- Actas Dermosifiliograficas Parte IDokumen8 halamanActas Dermosifiliograficas Parte Imariamv054Belum ada peringkat
- Therapeutic Potential of Stem Cells in Skin Repair and RegenerationDokumen13 halamanTherapeutic Potential of Stem Cells in Skin Repair and RegenerationTaro RahmatiaBelum ada peringkat
- The Chicken As A Model For Embryonic Development: M.G. Davey C. TickleDokumen9 halamanThe Chicken As A Model For Embryonic Development: M.G. Davey C. TickleUtari Septiana DewiBelum ada peringkat
- Cell BiolDokumen6 halamanCell BiolHamza AhmedBelum ada peringkat
- The Potential of Bovine Colostrum-Derived Exosomes To Repair Aged and Damaged Skin CellsDokumen15 halamanThe Potential of Bovine Colostrum-Derived Exosomes To Repair Aged and Damaged Skin CellsEduardo CastañedaBelum ada peringkat
- Follicular Development in The Human Infant OvaryDokumen13 halamanFollicular Development in The Human Infant OvaryRickySaptarshi SahaBelum ada peringkat
- ISSN 1679-9216 (Online)Dokumen19 halamanISSN 1679-9216 (Online)Rachel HayonBelum ada peringkat
- Bao Quan Mo Tinh HoanDokumen21 halamanBao Quan Mo Tinh Hoanhhhhhhhhhh0000Belum ada peringkat
- 2015-Urrego-Follicular Progesterone Concentrations and Messenger RNADokumen9 halaman2015-Urrego-Follicular Progesterone Concentrations and Messenger RNARodrigo UrregoBelum ada peringkat
- Synergy Between Genetic and Tissue Engineering: Creating An Artificial Salivary GlandDokumen6 halamanSynergy Between Genetic and Tissue Engineering: Creating An Artificial Salivary Glandammarstein89Belum ada peringkat
- Inv 14Dokumen8 halamanInv 14mrojasf.1996Belum ada peringkat
- Version On-Line ISSN 0717-9502: International Journal of MorphologyDokumen8 halamanVersion On-Line ISSN 0717-9502: International Journal of MorphologyMaría AlejandraBelum ada peringkat
- Kidney HistogenesisDokumen9 halamanKidney HistogenesisKshitija KasturiBelum ada peringkat
- Argan Vitalisation of Dermal Stem Cells For Skin Rejuvenation Personal Care 07 2011Dokumen3 halamanArgan Vitalisation of Dermal Stem Cells For Skin Rejuvenation Personal Care 07 2011Steve D'HamsBelum ada peringkat
- 2013 HongDokumen6 halaman2013 HongChloeee GBelum ada peringkat
- Ginther, 1998 Equine PregnancyDokumen32 halamanGinther, 1998 Equine PregnancyAdrian Ayala GutierrezBelum ada peringkat
- Epidermal Sphingolipids: Metabolism, Function, and Roles in Skin DisordersDokumen11 halamanEpidermal Sphingolipids: Metabolism, Function, and Roles in Skin DisordersymiyazyBelum ada peringkat
- Skin Integrated With Perfusable Vascular Channels On A ChipDokumen9 halamanSkin Integrated With Perfusable Vascular Channels On A ChipJ Ponmozhi IPSABelum ada peringkat
- Congenital Obstructive Hydrocephalus Revised 1 AutorecoveredDokumen7 halamanCongenital Obstructive Hydrocephalus Revised 1 Autorecoveredapi-652813198Belum ada peringkat
- Previews: Living in Peace: Host-Microbiota Mutualism in The SkinDokumen2 halamanPreviews: Living in Peace: Host-Microbiota Mutualism in The Skinblooom_00Belum ada peringkat
- Colostrogenesis - IgG1 Transcytosis Mechanisms 2014Dokumen15 halamanColostrogenesis - IgG1 Transcytosis Mechanisms 2014Jesús Enrique Turín VilcaBelum ada peringkat
- VishuDokumen4 halamanVishuVishuBelum ada peringkat
- Histopathological Study of Endometritis of The Cows: AbstractsDokumen6 halamanHistopathological Study of Endometritis of The Cows: Abstractsهدير عبدالمطلب حمزةBelum ada peringkat
- ID Pola Distribusi Mitokondria Sel Sel TrofDokumen10 halamanID Pola Distribusi Mitokondria Sel Sel TrofRahmad ArfandiBelum ada peringkat
- Experimental Dermatology - 2014 - Driskell - Defining Dermal Adipose TissueDokumen3 halamanExperimental Dermatology - 2014 - Driskell - Defining Dermal Adipose Tissueandre.ramosBelum ada peringkat
- Adaptationprocessoftheskingrafttovaginalmucosaafter MC IndoevaginoplastyDokumen5 halamanAdaptationprocessoftheskingrafttovaginalmucosaafter MC IndoevaginoplastyAnonymous UHnQSkxLBDBelum ada peringkat
- Cell RevolutionDokumen13 halamanCell RevolutionDream KyleBelum ada peringkat
- As Epidermal Stem Cells Age They Do Not Substantially Change Their CharacteristicsDokumen9 halamanAs Epidermal Stem Cells Age They Do Not Substantially Change Their CharacteristicsMichał PikułaBelum ada peringkat
- Bies 20137Dokumen15 halamanBies 20137Jerahmeel Sombilon GenillaBelum ada peringkat
- Experimental Design of A Culture Approach For Corneal Endothelial - 2020 - HeliDokumen8 halamanExperimental Design of A Culture Approach For Corneal Endothelial - 2020 - Helipo hongBelum ada peringkat
- Do Histologic Changes in The Skin-Lined Neovagina of Male-to-Female Transsexuals Really Occur?Dokumen4 halamanDo Histologic Changes in The Skin-Lined Neovagina of Male-to-Female Transsexuals Really Occur?momo1206Belum ada peringkat
- Lab 2-4 Formal ReportDokumen7 halamanLab 2-4 Formal ReportAbdullah AslamBelum ada peringkat
- Animals 11 01995Dokumen14 halamanAnimals 11 01995Jihan Natul Sa'diahBelum ada peringkat
- Age Related PermeabilityDokumen8 halamanAge Related Permeabilityabhijit_gothoskar6039Belum ada peringkat
- E I R H C:: Thical Ssues Egarding Uman Loning A Nursing PerspectiveDokumen18 halamanE I R H C:: Thical Ssues Egarding Uman Loning A Nursing PerspectiveNobin Kalayil BabuBelum ada peringkat
- Germline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofDokumen14 halamanGermline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofMariane Gabriela Cesar Ribeiro FerreiraBelum ada peringkat
- Impact of Different Antiseptics On Umbilical Cord Colonization and Cord Separation TimeDokumen6 halamanImpact of Different Antiseptics On Umbilical Cord Colonization and Cord Separation TimeFatma ElzaytBelum ada peringkat
- Stem Cell Dan Produknya Di UnnesDokumen56 halamanStem Cell Dan Produknya Di UnnesemyBelum ada peringkat
- The Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisDokumen10 halamanThe Mouse Epididymal Transcriptome: Transcriptional Profiling of Segmental Gene Expression in The EpididymisAhmad SolihinBelum ada peringkat
- Nuclear TransplantationDokumen6 halamanNuclear TransplantationDéépákBelum ada peringkat
- Der 073Dokumen9 halamanDer 073Sarrah AyuandariBelum ada peringkat
- Journal Collodion BabyDokumen5 halamanJournal Collodion BabyLidia MdBelum ada peringkat
- Information Networks in The Mammary Gland: Lothar Hennighausen and Gertraud W. RobinsonDokumen11 halamanInformation Networks in The Mammary Gland: Lothar Hennighausen and Gertraud W. RobinsonSueños De LunaBelum ada peringkat
- The Utility of Autologous Endometrial Coculture After Failed IVF A Report On 1194 Consecutive IVF Cycles From A Single InstitutionDokumen1 halamanThe Utility of Autologous Endometrial Coculture After Failed IVF A Report On 1194 Consecutive IVF Cycles From A Single InstitutionbbeeBelum ada peringkat
- Traumatic Brain Injury (TBI) - Definition and Pathophysiology - Overview, Epidemiology, Primary InjuryDokumen15 halamanTraumatic Brain Injury (TBI) - Definition and Pathophysiology - Overview, Epidemiology, Primary InjuryUtari Septiana DewiBelum ada peringkat
- Respiratorysystem ClozeDokumen1 halamanRespiratorysystem ClozeUtari Septiana DewiBelum ada peringkat
- Na Blog - CZ: Reproduction ClozeDokumen6 halamanNa Blog - CZ: Reproduction ClozeUtari Septiana DewiBelum ada peringkat
- Na Blog - CZ: ClozeDokumen6 halamanNa Blog - CZ: ClozeUtari Septiana DewiBelum ada peringkat
- Adapted From Observer - Guardian.co - UkDokumen1 halamanAdapted From Observer - Guardian.co - UkUtari Septiana DewiBelum ada peringkat
- PubMed Central, Table 1 - Arq Bras Cir Dig. 2014 Oct-Dec 27 (4) - 298-302. Doi - 10Dokumen1 halamanPubMed Central, Table 1 - Arq Bras Cir Dig. 2014 Oct-Dec 27 (4) - 298-302. Doi - 10Utari Septiana DewiBelum ada peringkat
- The Chicken As A Model For Embryonic Development: M.G. Davey C. TickleDokumen9 halamanThe Chicken As A Model For Embryonic Development: M.G. Davey C. TickleUtari Septiana DewiBelum ada peringkat
- Histology of Digestive SystemDokumen100 halamanHistology of Digestive SystemFadhila Putri Palupi100% (3)
- (Methods in Molecular Biology 1149) Alain Filloux, Juan-Luis Ramos (Eds.) - Pseudomonas Methods and Protocols-Humana Press (2014)Dokumen531 halaman(Methods in Molecular Biology 1149) Alain Filloux, Juan-Luis Ramos (Eds.) - Pseudomonas Methods and Protocols-Humana Press (2014)Muhammad ShakirBelum ada peringkat
- Benign and Malignant Tumors of Oral CavityDokumen56 halamanBenign and Malignant Tumors of Oral CavityPawan Kumar83% (6)
- NEOPLASIADokumen35 halamanNEOPLASIAnnBelum ada peringkat
- Reviewer PrelimDokumen19 halamanReviewer PrelimALPEREZ, GRACE LYN C.Belum ada peringkat
- Histopathological Study of Endometritis of The Cows: AbstractsDokumen6 halamanHistopathological Study of Endometritis of The Cows: Abstractsهدير عبدالمطلب حمزةBelum ada peringkat
- Reproductive SystemDokumen23 halamanReproductive SystemdrynwhylBelum ada peringkat
- Molecular Etiopathogenesis of Ameloblastoma CurrentDokumen5 halamanMolecular Etiopathogenesis of Ameloblastoma CurrentRiskha Febriani HapsariBelum ada peringkat
- Chapter 2 Cell Structure and Cell OrganisationDokumen75 halamanChapter 2 Cell Structure and Cell OrganisationleelijyeBelum ada peringkat
- Summary Assessment HISTO LABDokumen8 halamanSummary Assessment HISTO LABARAH DE OCAMPOBelum ada peringkat
- Composed of Closely Aggregated Polyhedral Cells With Strong Adhesion To One AnotherDokumen3 halamanComposed of Closely Aggregated Polyhedral Cells With Strong Adhesion To One AnotherWinnie Gay UanangBelum ada peringkat
- Histology EpitheliumDokumen18 halamanHistology EpitheliumMarina TeahariBelum ada peringkat
- Webquest - HistologyDokumen6 halamanWebquest - HistologyicdjpBelum ada peringkat
- (Cancer and Vitamin C) Ebook PDF by (Linus Pauling), Cameron, Ewan Pauling, Linus, 1901-1994, JDokumen260 halaman(Cancer and Vitamin C) Ebook PDF by (Linus Pauling), Cameron, Ewan Pauling, Linus, 1901-1994, JAnonymous gwFqQcnaX100% (2)
- NTSE Biology Questions: Chapter(s) : Cell-Fundamental Unit of Life & TissuesDokumen60 halamanNTSE Biology Questions: Chapter(s) : Cell-Fundamental Unit of Life & Tissuesss SBelum ada peringkat
- Histo CompilationDokumen217 halamanHisto CompilationNico LokoBelum ada peringkat
- 2 - Anatomy and Histology of Female Genital TractDokumen26 halaman2 - Anatomy and Histology of Female Genital TractHervis FantiniBelum ada peringkat
- BJS 16-3 PDFDokumen59 halamanBJS 16-3 PDFJhonatan Paredes VásquezBelum ada peringkat
- Epithelial Tissues REVIEWERDokumen16 halamanEpithelial Tissues REVIEWERClyde BaltazarBelum ada peringkat
- Dentigerous CystDokumen25 halamanDentigerous CystDr. Deepanshi SutariaBelum ada peringkat
- Neville, Oral and Maxillofacial Pathology, 2nd EdDokumen857 halamanNeville, Oral and Maxillofacial Pathology, 2nd EdPatriciaSepúlvedaCruz67% (3)
- 3.mechanical InjuryDokumen127 halaman3.mechanical InjuryARIF-UR-REHMAN100% (2)
- DR - Moslim Obs & Gyn Answers GuideDokumen124 halamanDR - Moslim Obs & Gyn Answers GuideMandark0009100% (5)
- 3.skin PathologyDokumen38 halaman3.skin PathologyFaisal MehboobBelum ada peringkat
- 10 Squamouspapilloma-ReportoftwocasesDokumen7 halaman10 Squamouspapilloma-ReportoftwocasesAyik DarkerThan BlackBelum ada peringkat
- Screenshot 2023-12-10 at 10.38.50 AMDokumen75 halamanScreenshot 2023-12-10 at 10.38.50 AMmizare29gBelum ada peringkat
- Lab 5 Tissues - Lab ReportDokumen5 halamanLab 5 Tissues - Lab ReportbrittBelum ada peringkat
- Testicular TumorsDokumen14 halamanTesticular TumorsJose SirittBelum ada peringkat
- Modul Final Biologi Form 4 Fasa 1 2021Dokumen60 halamanModul Final Biologi Form 4 Fasa 1 2021Manmohon Kaur100% (1)
- Master File - ChronicDokumen671 halamanMaster File - ChronicsubhojitnayekBelum ada peringkat