Anda mungkin juga menyukai
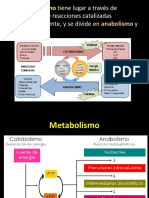
- 12 MetabolismoDokumen80 halaman12 MetabolismoJean Paul E Mendez CotaBelum ada peringkat
- Respiración CelularDokumen25 halamanRespiración Celularjessisegura365Belum ada peringkat
- Guia Teorica Unidad 4 y Mapa ConceptualDokumen10 halamanGuia Teorica Unidad 4 y Mapa ConceptualJavier F Rosero MBelum ada peringkat
- Tema 7Dokumen34 halamanTema 7daniel salazarBelum ada peringkat
- CATABOLISMODokumen13 halamanCATABOLISMOsilvanaBelum ada peringkat
- F4METABOLISMOMICROBIANODokumen20 halamanF4METABOLISMOMICROBIANOSheylla M RamosBelum ada peringkat
- Nutrición y Metabolismo BacterianoDokumen35 halamanNutrición y Metabolismo BacterianoJuan Camilo Cardona Salazar100% (1)
- Nutrición, Metabolismo y Fisiología BacterianaDokumen36 halamanNutrición, Metabolismo y Fisiología Bacterianapepito perezBelum ada peringkat
- 12 MetabolismoDokumen92 halaman12 Metabolismobresler_linBelum ada peringkat
- Clase RespiraciónDokumen27 halamanClase RespiraciónRUBEN JOSUE MIGUEL BASILIO MIRANDABelum ada peringkat
- Formato Tarea 3 Ejercicio IndividualDokumen4 halamanFormato Tarea 3 Ejercicio Individualliceth gutierrezBelum ada peringkat
- Introducción Al Metabolismo y Bioenergética.Dokumen28 halamanIntroducción Al Metabolismo y Bioenergética.Emil DiazBelum ada peringkat
- Unidad 7, Introduccion Metabolismo CelularDokumen61 halamanUnidad 7, Introduccion Metabolismo CelularignacioBelum ada peringkat
- Clase 5 - Metabolismo PrimarioDokumen52 halamanClase 5 - Metabolismo PrimarioLEMONBelum ada peringkat
- Respiracion Aerobia y AnaerobiaDokumen21 halamanRespiracion Aerobia y AnaerobiaBetsi Galeana100% (1)
- Utilizacion de Energia Por Los MicroorganismosDokumen47 halamanUtilizacion de Energia Por Los MicroorganismosBrendaBelum ada peringkat
- Fases de La Respiración CelularDokumen2 halamanFases de La Respiración CelularNoha VanegasBelum ada peringkat
- Metabolismo BacterianoDokumen80 halamanMetabolismo BacterianokatherynBelum ada peringkat
- Catabolismo y AnabolismoDokumen10 halamanCatabolismo y AnabolismoPilar Tolos EscorihuelaBelum ada peringkat
- Respiracion CelularDokumen72 halamanRespiracion CelularSheila Katheryn Paucar Ancco100% (1)
- Resumen Exposicion FermentaciónDokumen3 halamanResumen Exposicion Fermentaciónanyel bermudezBelum ada peringkat
- 8a Ferment 23Dokumen20 halaman8a Ferment 23Lineth BCBelum ada peringkat
- Semana 10. Obtención de Energía UPNDokumen35 halamanSemana 10. Obtención de Energía UPNcarmen marquinaBelum ada peringkat
- La Mitocondria y La Glucolisis 1Dokumen60 halamanLa Mitocondria y La Glucolisis 1Henry Raul VillatoroBelum ada peringkat
- MetabolismoDokumen65 halamanMetabolismoItzel PergonBelum ada peringkat
- Biotransformacion de La EnergíaDokumen82 halamanBiotransformacion de La EnergíaLia Miranda Guillen100% (3)
- Cata y AnaDokumen3 halamanCata y AnaDiana LauraBelum ada peringkat
- CATABOLISMO (Aerobio y Anaerobio)Dokumen66 halamanCATABOLISMO (Aerobio y Anaerobio)Elizabetн Essel Cordilia Middleford50% (2)
- Clase N°03 FermentacionesDokumen38 halamanClase N°03 FermentacionesJeanpierre Quiroz100% (1)
- 8) Met Bact DR BarlettaDokumen18 halaman8) Met Bact DR BarlettaZósimo SuasnabarBelum ada peringkat
- El Proceso de Fermentacion y Los Microoganismos...Dokumen10 halamanEl Proceso de Fermentacion y Los Microoganismos...Valentina Ruiz RamírezBelum ada peringkat
- MetabolismoDokumen67 halamanMetabolismoSam Alarcon MartinezBelum ada peringkat
- Metabolismo BacterianoDokumen8 halamanMetabolismo BacterianoArévalo100% (1)
- 2 Metabolismo MicrobianoDokumen101 halaman2 Metabolismo MicrobianoJefferson OchoaBelum ada peringkat
- Clase 09 - Respiracion CelularDokumen34 halamanClase 09 - Respiracion CelularLivia NinaBelum ada peringkat
- Clase 9. FermentaciónDokumen18 halamanClase 9. FermentaciónDaniela Estrella AracenaBelum ada peringkat
- Metabolismo CelularDokumen20 halamanMetabolismo CelularYanira Castellano EstevezBelum ada peringkat
- Guia BioquimicaDokumen7 halamanGuia BioquimicaSamanta ZariñanBelum ada peringkat
- Metabolismo de MicroorganismosDokumen46 halamanMetabolismo de MicroorganismosJOHANA BECERRABelum ada peringkat
- Catabolismo PDFDokumen77 halamanCatabolismo PDFAnny SnyBelum ada peringkat
- METABOLISMODokumen16 halamanMETABOLISMODavid VelascoBelum ada peringkat
- CATABOLISMODokumen8 halamanCATABOLISMOMerry Peralta VillanuevaBelum ada peringkat
- Concepto de FermentaciónDokumen62 halamanConcepto de FermentaciónneothonyBelum ada peringkat
- Ciclo Del Ácido CítricoDokumen44 halamanCiclo Del Ácido CítricoJm AlmarazBelum ada peringkat
- Metabolismo y Respiración CelularDokumen26 halamanMetabolismo y Respiración CelularHerbert JaimesBelum ada peringkat
- QuimioheterótrofosDokumen11 halamanQuimioheterótrofosMiguel Sanchez CastilloBelum ada peringkat
- Metabolismo, Actividad EnzimaticaDokumen5 halamanMetabolismo, Actividad EnzimaticaAstrid GonzalezBelum ada peringkat
- Respiracion CelularDokumen34 halamanRespiracion CelularMaria Jose ROBelum ada peringkat
- FermentaciónDokumen42 halamanFermentaciónChava HernandezBelum ada peringkat
- Catabolismo y AnabolismoDokumen25 halamanCatabolismo y AnabolismoMiguel ChávezBelum ada peringkat
- Respiracion CelularDokumen45 halamanRespiracion CelularAlelyBelum ada peringkat
- 2º Bachillerato Biología TEMA 11 LOMCEDokumen44 halaman2º Bachillerato Biología TEMA 11 LOMCEIzumi MinBelum ada peringkat
- Fermentacion PDFDokumen23 halamanFermentacion PDFKATYABelum ada peringkat
- Metabolismo CelularDokumen10 halamanMetabolismo Celularalexandra romeroBelum ada peringkat
- Tema 10.1. Metabolismo: Catabolismo: 1.-Definición Y Características GeneralesDokumen7 halamanTema 10.1. Metabolismo: Catabolismo: 1.-Definición Y Características GeneralesManuela GaleanoBelum ada peringkat
- Metabolismo MicrobianoDokumen68 halamanMetabolismo MicrobianoKary SantoméBelum ada peringkat
- LitotrofismoDokumen44 halamanLitotrofismoMaria Del Pilar Vilchez MolinaBelum ada peringkat
- Tarea V Biologia y Su Lab... Emei BelloDokumen32 halamanTarea V Biologia y Su Lab... Emei BelloESMARLIS BELLO JOSE100% (1)
- Ejercicios Flotaciòn 1-8-9Dokumen6 halamanEjercicios Flotaciòn 1-8-9J Quispe Montañez100% (1)
- Metalurgia 1 Tarea 3.1Dokumen3 halamanMetalurgia 1 Tarea 3.1J Quispe MontañezBelum ada peringkat
- Problemas de Flotación de MineralesDokumen57 halamanProblemas de Flotación de MineralesJ Quispe Montañez100% (8)
- Problemas Flotacion 1Dokumen13 halamanProblemas Flotacion 1J Quispe Montañez100% (1)
- Piedra LajaDokumen7 halamanPiedra LajaJ Quispe MontañezBelum ada peringkat
- Metalurgia 1 Tarea ProblemasDokumen6 halamanMetalurgia 1 Tarea ProblemasJ Quispe Montañez0% (1)
- Metodo de La SecanteDokumen6 halamanMetodo de La SecanteJ Quispe MontañezBelum ada peringkat
- Problemas para ExamenDokumen1 halamanProblemas para ExamenJ Quispe MontañezBelum ada peringkat
- Examen 1A para La CasaDokumen2 halamanExamen 1A para La CasaJ Quispe MontañezBelum ada peringkat
- VariablesDokumen6 halamanVariablesJ Quispe MontañezBelum ada peringkat
- Plasticos Biodegradables Alm1Dokumen22 halamanPlasticos Biodegradables Alm1J Quispe MontañezBelum ada peringkat
- Ejercicios Resueltos de FenomenosDokumen10 halamanEjercicios Resueltos de FenomenosJ Quispe MontañezBelum ada peringkat
- BioreactoresDokumen56 halamanBioreactoresJ Quispe MontañezBelum ada peringkat
- Procariotas 2 PDFDokumen59 halamanProcariotas 2 PDFJ Quispe MontañezBelum ada peringkat
- Peste NegraDokumen7 halamanPeste NegraJulian ZuluagaBelum ada peringkat
- CROMOBLASTOMICOSISDokumen15 halamanCROMOBLASTOMICOSISJuan A. JambBelum ada peringkat
- Celulas GlialesDokumen5 halamanCelulas GlialesNorma Luisa L. EBelum ada peringkat
- Escritura en EspejoDokumen12 halamanEscritura en EspejoFreddy Espejo100% (1)
- Alergia A MedicamentosDokumen38 halamanAlergia A MedicamentosAntonella Castro CabreraBelum ada peringkat
- Diagnóstico Molecular de Los Déficit de 21-Hidroxilasa y Su Correlación Con El Fenotipo - Endocrinología y NutriciónDokumen12 halamanDiagnóstico Molecular de Los Déficit de 21-Hidroxilasa y Su Correlación Con El Fenotipo - Endocrinología y NutriciónJosue BarralBelum ada peringkat
- Hallazgos Patológicos en Radiografías Panorámicas Previas Al Tratamiento de Ortodoncia en El La Clínica Odontológica de La Uap en El Año 2018Dokumen2 halamanHallazgos Patológicos en Radiografías Panorámicas Previas Al Tratamiento de Ortodoncia en El La Clínica Odontológica de La Uap en El Año 2018Eliza EspinozaBelum ada peringkat
- 10 Maneras de Utilizar El Aloe Vera o SábilaDokumen6 halaman10 Maneras de Utilizar El Aloe Vera o SábilaAnonymous HmRlPGIBelum ada peringkat
- Anomalias Congenitas Del Aparato Genital FemeninoDokumen56 halamanAnomalias Congenitas Del Aparato Genital Femeninodarwin2175975% (4)
- Anatomia Crecimiento Del VelloDokumen4 halamanAnatomia Crecimiento Del VelloAdriana OmañaBelum ada peringkat
- Efecto de La Temperatura Sustancias en La Funcion Cardiaca en AnfibiosDokumen8 halamanEfecto de La Temperatura Sustancias en La Funcion Cardiaca en AnfibiosAngelManriqueCristobalBelum ada peringkat
- I. Control Del Estado NeurológicoDokumen56 halamanI. Control Del Estado Neurológicokaren1garcia_2Belum ada peringkat
- LARINGITISDokumen2 halamanLARINGITISCarlos GuidoBelum ada peringkat
- Charla MayDokumen5 halamanCharla MayMay Larrea Cubas0% (1)
- Triptico de EmocionDokumen2 halamanTriptico de EmocionSoledad CastilloBelum ada peringkat
- Informacion de Terio Expo PraDokumen2 halamanInformacion de Terio Expo PraKingdoms OrouteBelum ada peringkat
- DSM-5 Cómo Hacer El Reporte PDFDokumen1 halamanDSM-5 Cómo Hacer El Reporte PDFAndr Greek100% (2)
- 3 Etiologia de Los Trastornos Del DesarrolloDokumen26 halaman3 Etiologia de Los Trastornos Del DesarrolloCaro LiBelum ada peringkat
- Estos Son Los 5 Venenos Blancos Que Deberías Evitar en Tu Consumo DiarioDokumen3 halamanEstos Son Los 5 Venenos Blancos Que Deberías Evitar en Tu Consumo DiarioJanela Arias PeñaBelum ada peringkat
- NEUROCIENCIASDokumen6 halamanNEUROCIENCIASHuaman Ramos IsamarBelum ada peringkat
- Anemia Por Deficiencia de HierroDokumen6 halamanAnemia Por Deficiencia de HierromayeyesBelum ada peringkat
- Clasificacion DisfemiaDokumen3 halamanClasificacion Disfemiaveronica mashingash petsaBelum ada peringkat
- Teoria NeurologicaDokumen5 halamanTeoria NeurologicaKarl_KAst_4993Belum ada peringkat
- Comparación Manual RotatorioDokumen10 halamanComparación Manual RotatorioJosé ChampetBelum ada peringkat
- GPC11 DiabetesDokumen14 halamanGPC11 DiabetesGilberto Juvencio Solano Aparicio100% (2)
- Triptico EscoliosisDokumen3 halamanTriptico EscoliosisAriel CristianBelum ada peringkat
- Alteración de Mecanismos FibrinolíticosDokumen35 halamanAlteración de Mecanismos FibrinolíticosMarcelo Eduardo Andrade CastroBelum ada peringkat
- Hormonas Peptidicas y Hormonas de CrecimientoDokumen41 halamanHormonas Peptidicas y Hormonas de CrecimientoCarlosBahenaBelum ada peringkat
- Lilloa 45 - Suplemento - InteriorDokumen204 halamanLilloa 45 - Suplemento - InteriorFundacionLillo0% (1)
- GabaDokumen6 halamanGabaRenee DayBelum ada peringkat