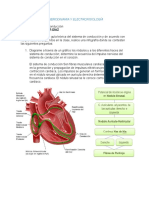
Anda mungkin juga menyukai
- Guía de Estudio 3Dokumen12 halamanGuía de Estudio 3Debora RuizBelum ada peringkat
- Interpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasDari EverandInterpretación del ECG: Una Guía Práctica e Intuitiva para Aprender a Leer el ECG y Diagnosticar y Tratar ArritmiasBelum ada peringkat
- E Investigación Biomédica - Electrocardiografía (ECG) - Dalcame Grupo DDokumen1 halamanE Investigación Biomédica - Electrocardiografía (ECG) - Dalcame Grupo Ddiana lucia serrano gomezBelum ada peringkat
- Excitoconductor y EKGDokumen16 halamanExcitoconductor y EKGAlbert FigueredoBelum ada peringkat
- 2 - Hipertrofia y Dilatación CardíacasDokumen45 halaman2 - Hipertrofia y Dilatación Cardíacas10280457Belum ada peringkat
- Fisiologia Del Corazon T 2Dokumen47 halamanFisiologia Del Corazon T 2Milagro Adriana Torres MondragónBelum ada peringkat
- 07 - Propiedades Del Músculo CardiacoDokumen11 halaman07 - Propiedades Del Músculo CardiacoAlejandra HernandezBelum ada peringkat
- Monografia Terminos FrecuentesDokumen18 halamanMonografia Terminos FrecuentesJulissa Poma CerronBelum ada peringkat
- Desfibrilador CardiacoDokumen46 halamanDesfibrilador CardiacoJesus Resta NavarroBelum ada peringkat
- 1.1. - Ekg 2021Dokumen64 halaman1.1. - Ekg 2021Diana CarstairsBelum ada peringkat
- Guia #8 - Electrocardiografía (Ecg)Dokumen43 halamanGuia #8 - Electrocardiografía (Ecg)Deisy BedoyaBelum ada peringkat
- 02 Actividad Electrica Del Corazon - EcgDokumen31 halaman02 Actividad Electrica Del Corazon - Ecgandres felipe montoya bolañosBelum ada peringkat
- CardiacoDokumen9 halamanCardiacodin 1938Belum ada peringkat
- (Estefany) Propiedades Electrofisiológicas Del Miocardio Contráctil y EspecializadoDokumen19 halaman(Estefany) Propiedades Electrofisiológicas Del Miocardio Contráctil y EspecializadoPIKABelum ada peringkat
- Electrofisiologia CardiacaDokumen3 halamanElectrofisiologia CardiacaMaria Eugenia GarciaBelum ada peringkat
- ElectrocardiogramaDokumen4 halamanElectrocardiogramaHector Josue Ramos CeronBelum ada peringkat
- Medidas de Señales Bioelectricas, Clase de La Universidad Nacional Experimental FMDokumen43 halamanMedidas de Señales Bioelectricas, Clase de La Universidad Nacional Experimental FMLuis LopezBelum ada peringkat
- Electrocardiografia BasicaDokumen43 halamanElectrocardiografia BasicaNana Rodriguez0% (1)
- Apunte Fisiología 3Dokumen4 halamanApunte Fisiología 3Lizeth ChicoBelum ada peringkat
- Actividad Eléctrica y Contractil Del CorazónDokumen7 halamanActividad Eléctrica y Contractil Del CorazónTrustter RomeroBelum ada peringkat
- Introductorio CardiovascularDokumen20 halamanIntroductorio CardiovascularViri MartínezBelum ada peringkat
- Electrocardiograma 01Dokumen15 halamanElectrocardiograma 01Yasirys Mariell PichardoBelum ada peringkat
- Practica de Laboratorio N0. 7Dokumen15 halamanPractica de Laboratorio N0. 7Edgar Osveli Temaj MacarioBelum ada peringkat
- CONDUCTIVIDADDokumen17 halamanCONDUCTIVIDADSurian CasasBelum ada peringkat
- CARDIOVASCULAR - CompressedDokumen10 halamanCARDIOVASCULAR - CompressedFrancisco Oliva EntebiBelum ada peringkat
- Teoría-Función Cardiovascular - MEHU 632Dokumen24 halamanTeoría-Función Cardiovascular - MEHU 632Sara ParedesBelum ada peringkat
- Cómo Leer Un ElectrocardiogramaDokumen17 halamanCómo Leer Un ElectrocardiogramaSusan Rivera ZabalaBelum ada peringkat
- AntiarritmiasDokumen40 halamanAntiarritmiasJosé Emilio Magaña TejadaBelum ada peringkat
- Tema 3.PROPIEDADES ELECTROFISIOLÓGICAS DEL MIOCARDIO CONTRÁCTIL Y ESPECIALIZADODokumen7 halamanTema 3.PROPIEDADES ELECTROFISIOLÓGICAS DEL MIOCARDIO CONTRÁCTIL Y ESPECIALIZADOyuri goitiaBelum ada peringkat
- PRACTICADokumen8 halamanPRACTICAmileidy inga romeroBelum ada peringkat
- Sistema Cardiovascular 2 - JEsús GonzálezDokumen13 halamanSistema Cardiovascular 2 - JEsús GonzálezJesús GonzálezBelum ada peringkat
- Corriente Eléctrica (FormatoDokumen37 halamanCorriente Eléctrica (FormatoGrecia NizamaBelum ada peringkat
- Propiedades Fisiológicas Del Músculo CardiacoDokumen14 halamanPropiedades Fisiológicas Del Músculo CardiacoMaría Belén MareskyBelum ada peringkat
- CardioooDokumen5 halamanCardioooapi-500143767Belum ada peringkat
- Propiedades Electricas Del CorazonDokumen4 halamanPropiedades Electricas Del CorazonRichard Armijo GarridoBelum ada peringkat
- Fisiología Del Corazón Guyton (Integración Eléctrica, Mecánica, Hemodinámica y Sonora)Dokumen14 halamanFisiología Del Corazón Guyton (Integración Eléctrica, Mecánica, Hemodinámica y Sonora)Gianfranco Tovar CalderónBelum ada peringkat
- Aparato Cardiovascular 1T1Dokumen41 halamanAparato Cardiovascular 1T1Marycielo LmBelum ada peringkat
- Grupo C - Actividad 2cDokumen9 halamanGrupo C - Actividad 2cmicacampos.baBelum ada peringkat
- Taller Lab Tutor ECG (1) OLGA (1) 234Dokumen13 halamanTaller Lab Tutor ECG (1) OLGA (1) 234Kane HookerBelum ada peringkat
- Paper-Ecg - Quispe Navarro JordanDokumen6 halamanPaper-Ecg - Quispe Navarro JordanMichaell Jordan Jonathan Quispe NavarroBelum ada peringkat
- Electrocardiograma Fases de Repolarizacion MiocardicaDokumen10 halamanElectrocardiograma Fases de Repolarizacion MiocardicaGrissel Bolaños SuarezBelum ada peringkat
- Biofisica CardíacaDokumen28 halamanBiofisica CardíacaMarcos Días100% (1)
- Electrocardiograma Normal y Patológico - Medicina InternaDokumen35 halamanElectrocardiograma Normal y Patológico - Medicina InternaPedro HermosoBelum ada peringkat
- Fisiologia CardiovascularDokumen5 halamanFisiologia CardiovascularYinna Colina LugoBelum ada peringkat
- Manual Electrocardiografia Basica e Interpretacion de ArritmiasDokumen35 halamanManual Electrocardiografia Basica e Interpretacion de ArritmiasAdrian LindemannBelum ada peringkat
- Fisiología Cardiovascular2017DagrosaDokumen76 halamanFisiología Cardiovascular2017DagrosaLautaro PreteBelum ada peringkat
- ElectrocardiogramaDokumen11 halamanElectrocardiogramaJaime AguirreBelum ada peringkat
- Electrofisiologia PDFDokumen90 halamanElectrofisiologia PDFFabiola NorabuenaBelum ada peringkat
- Fisiologia CardiacaDokumen11 halamanFisiologia CardiacaCarol IsidroBelum ada peringkat
- Nodulo SinusalDokumen5 halamanNodulo SinusalDaniel Noriega MezaBelum ada peringkat
- Marcapasos y DesfibriladoresDokumen7 halamanMarcapasos y Desfibriladorescristhian salasBelum ada peringkat
- Mod-1 Tema-2 ElectrofisiologíaDokumen8 halamanMod-1 Tema-2 Electrofisiologíadieguitotapia77Belum ada peringkat
- Unidad 5 - Anatomia y Fisiologia CardiacaDokumen4 halamanUnidad 5 - Anatomia y Fisiologia CardiacaAnonymous c3HKuIBelum ada peringkat
- Electrofisiologia Cardiaca Junio 2007Dokumen36 halamanElectrofisiologia Cardiaca Junio 2007Diana Sánchez CuentasBelum ada peringkat
- El ElectrocardiogramaDokumen2 halamanEl ElectrocardiogramaCristian Fernando Esquivel CerpaBelum ada peringkat
- Clase 1 Electrofisiologia CardiacaDokumen68 halamanClase 1 Electrofisiologia CardiacaCarlos fenixBelum ada peringkat
- Electrocardiografo Material de ApoyoDokumen34 halamanElectrocardiografo Material de ApoyoRonny Juseph Garcia NiñoBelum ada peringkat
- Informe de EKGDokumen16 halamanInforme de EKGAnthony PalaciosBelum ada peringkat
- Potencial de Membrana y CardiovasculaDokumen7 halamanPotencial de Membrana y CardiovasculaJacki ServianBelum ada peringkat
- Mec Musculo EsqDokumen2 halamanMec Musculo EsqDavid ValdésBelum ada peringkat
- AnticuerposDokumen30 halamanAnticuerposDavid ValdésBelum ada peringkat
- Legislación Productos CosméticosDokumen5 halamanLegislación Productos CosméticosRamón ZazuetaBelum ada peringkat
- El Arco ReflejoADokumen7 halamanEl Arco ReflejoAIsabel Jessica Támara GuzmánBelum ada peringkat
- Adsorcion 1Dokumen4 halamanAdsorcion 1David ValdésBelum ada peringkat
- Hormona Del Crecimiento de Fisio HumanaDokumen11 halamanHormona Del Crecimiento de Fisio HumanaDavid ValdésBelum ada peringkat
- GLCDokumen1 halamanGLCDavid ValdésBelum ada peringkat
- Extracción de Nicotina de FarmoDokumen14 halamanExtracción de Nicotina de FarmoDavid ValdésBelum ada peringkat
- Arco ReflejoDokumen7 halamanArco ReflejoDavid ValdésBelum ada peringkat
- Anexo Soluciones ParenteralesDokumen3 halamanAnexo Soluciones ParenteralesDanny GuevaraBelum ada peringkat
- Sintesisyreaccionesindolybenzofurano 11063Dokumen111 halamanSintesisyreaccionesindolybenzofurano 11063Mario Leonardo Guevara EscalanteBelum ada peringkat
- Escala ECG PDFDokumen1 halamanEscala ECG PDFDavid ValdésBelum ada peringkat
- Anexo Soluciones ParenteralesDokumen3 halamanAnexo Soluciones ParenteralesDanny GuevaraBelum ada peringkat
- Tierra RojaDokumen5 halamanTierra RojaDavid ValdésBelum ada peringkat
- Insta Laci OnesDokumen8 halamanInsta Laci OnesDavid ValdésBelum ada peringkat
- P. S. PI ProteínasDokumen2 halamanP. S. PI ProteínasDavid ValdésBelum ada peringkat
- BotanicaDokumen7 halamanBotanicaDavid ValdésBelum ada peringkat
- Aplic. Prác. E. MurosDokumen1 halamanAplic. Prác. E. MurosDavid ValdésBelum ada peringkat
- SIMULACRODokumen4 halamanSIMULACRODavid ValdésBelum ada peringkat
- Deuda de OxigenoDokumen5 halamanDeuda de OxigenoCesarBelum ada peringkat
- Electrocardiografía: Biblioteca Medica VirutalDokumen9 halamanElectrocardiografía: Biblioteca Medica Virutaljessica perezBelum ada peringkat
- Tema 9. Metabolismo 2022Dokumen59 halamanTema 9. Metabolismo 2022Franco GonzálezBelum ada peringkat
- GASOMETRÍADokumen23 halamanGASOMETRÍAMarco Antonio De Jesus CorreaBelum ada peringkat
- Transfusión de HemocomponentesDokumen40 halamanTransfusión de HemocomponentesGladys Selene100% (1)
- 03 03-Arritmias PDFDokumen14 halaman03 03-Arritmias PDFnancybernalBelum ada peringkat
- Desarrollo Fetal y Cambios de La GestanteDokumen4 halamanDesarrollo Fetal y Cambios de La GestanteCarolina VegaBelum ada peringkat
- Qué Son Las NeuronasDokumen6 halamanQué Son Las NeuronasMaria Jose Toscano GonzálezBelum ada peringkat
- Uvm PRÁCTICA #11Dokumen10 halamanUvm PRÁCTICA #11Katherine Manriquez TrenadoBelum ada peringkat
- Signos Vitales 25-05-2022Dokumen49 halamanSignos Vitales 25-05-2022Kndla NajeraBelum ada peringkat
- Triptico El Tejido EpitelialDokumen2 halamanTriptico El Tejido EpitelialDIEGO LEONARDO ROMERO ROMEROBelum ada peringkat
- Emergencia Hipertensiva en Uci DesarrolloDokumen8 halamanEmergencia Hipertensiva en Uci DesarrolloGema Ojeda RodríguezBelum ada peringkat
- Biologia 8Dokumen78 halamanBiologia 8Santos Canahuire CondoriBelum ada peringkat
- 3°a Practica Sistema CirculatorioDokumen4 halaman3°a Practica Sistema CirculatorioAlexsandra Gutierrez PeñaBelum ada peringkat
- Exposición Cadena RespiratoriaDokumen5 halamanExposición Cadena RespiratoriaCarlos Manuel Narvaez CifuentesBelum ada peringkat
- TROMBECTOMÍADokumen10 halamanTROMBECTOMÍAJennifer100% (1)
- Sistemas de Conduccion Vid 2 PDFDokumen76 halamanSistemas de Conduccion Vid 2 PDFRita MelisaBelum ada peringkat
- Tejido Sanguineo PDFDokumen9 halamanTejido Sanguineo PDFLuckiBelum ada peringkat
- Ejercicio AnaeróbicoDokumen3 halamanEjercicio AnaeróbicoRosita Rosvic100% (1)
- Alteraciones Del Nivel de Conciencia y ComaDokumen51 halamanAlteraciones Del Nivel de Conciencia y ComaLizandra Silva AlonsoBelum ada peringkat
- Cardiovascular en El DeporteDokumen320 halamanCardiovascular en El DeporteFernando DE LA Torre0% (1)
- Evaluación AntropométricaDokumen5 halamanEvaluación AntropométricaSebastián Robledo MarínBelum ada peringkat
- Metabolismo CelularDokumen3 halamanMetabolismo CelularAnaBelum ada peringkat
- Unidad 45 Semiología Del Sistema NerviosoDokumen14 halamanUnidad 45 Semiología Del Sistema NerviosoDhery Prieto RuizBelum ada peringkat
- Inflamación y Reparación PatologíaDokumen19 halamanInflamación y Reparación PatologíaLuliBelum ada peringkat
- Sistema Respiratorio 4°Dokumen5 halamanSistema Respiratorio 4°Raquel Acevedo JaimesBelum ada peringkat
- Semiología de La Disnea. 2021Dokumen37 halamanSemiología de La Disnea. 2021Adela Garcete100% (1)
- Tema Neurotransmisores - ExposiciónDokumen19 halamanTema Neurotransmisores - ExposiciónIngTermineitorQuispeBelum ada peringkat
- Tema 2Dokumen13 halamanTema 2Noelia100% (1)
- Transporte de O2 y Co2 Vesalius MedicDokumen19 halamanTransporte de O2 y Co2 Vesalius MedicnoebladimirBelum ada peringkat