Anda mungkin juga menyukai
- PTERIDOPHYTA FYBSc NotesDokumen12 halamanPTERIDOPHYTA FYBSc Notesvishcrime100% (4)
- Pteridophytes FYBSc Notes FinalDokumen16 halamanPteridophytes FYBSc Notes Finalvishcrime83% (12)
- Morphology of Flowering PlantsDokumen24 halamanMorphology of Flowering PlantsNalla Raghuram ChowdaryBelum ada peringkat
- F.y.b.sc - Botany Sem. I, IIDokumen15 halamanF.y.b.sc - Botany Sem. I, IIPraveen Kumar100% (1)
- CBSE Quick Revision Notes CBSE Class-11 Biology Chapter-11 Transport in PlantsDokumen10 halamanCBSE Quick Revision Notes CBSE Class-11 Biology Chapter-11 Transport in PlantsPriyanshu BhadanaBelum ada peringkat
- Class 11th Morphology of Flowering Plants SolutionsDokumen13 halamanClass 11th Morphology of Flowering Plants SolutionsMufeedAliBelum ada peringkat
- Anatomy of Flowering Plants - Shobhit NirwanDokumen15 halamanAnatomy of Flowering Plants - Shobhit NirwanPrakhar SrivastavaBelum ada peringkat
- Performance Enhancement Biology Basics Part 1Dokumen6 halamanPerformance Enhancement Biology Basics Part 1Tera Bap50% (2)
- MORPHOLOGY OF FLOWERING PLANTSDokumen15 halamanMORPHOLOGY OF FLOWERING PLANTSAanchal Pandey100% (1)
- Palaeobotany: Types Nomenclature of FossilsDokumen21 halamanPalaeobotany: Types Nomenclature of FossilsPawan KatiyarBelum ada peringkat
- Morphological Features of CycasDokumen41 halamanMorphological Features of CycasHamid JoiyaBelum ada peringkat
- Key Notes On Chemical Bonding: Crash Course For NEET 2021Dokumen6 halamanKey Notes On Chemical Bonding: Crash Course For NEET 2021thorBelum ada peringkat
- NEET Biology Crash Course Key NotesDokumen5 halamanNEET Biology Crash Course Key NotesHerick RobinBelum ada peringkat
- Ecology: Botany by DR Geetendra Mbbs MD @Dokumen35 halamanEcology: Botany by DR Geetendra Mbbs MD @Débàshis Dash100% (1)
- Exp No 7 Meiosis in Onion Bud CellDokumen3 halamanExp No 7 Meiosis in Onion Bud CellShadab AhmadBelum ada peringkat
- Division Bryophyta PDFDokumen29 halamanDivision Bryophyta PDFPee BeeBelum ada peringkat
- Morphology of Flowering Plants: Mind MapsDokumen4 halamanMorphology of Flowering Plants: Mind MapsRiya Bhardwaj100% (1)
- Class Xii BiologyDokumen63 halamanClass Xii Biologyairs primeBelum ada peringkat
- Pollen: Jump To Navigationjump To SearchDokumen19 halamanPollen: Jump To Navigationjump To Searchjessito2100% (2)
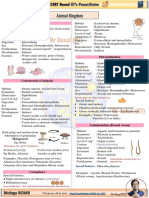
- Animal Kingdom PowernotesDokumen3 halamanAnimal Kingdom PowernotesSushmit SrivastavaBelum ada peringkat
- The Living World: Mind MapDokumen12 halamanThe Living World: Mind MapPrince100% (1)
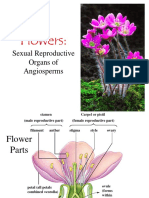
- Flowers: Sexual Reproduction in AngiospermsDokumen106 halamanFlowers: Sexual Reproduction in AngiospermsAlfonso PlantillaBelum ada peringkat
- Principles of Inheritance and Variation-NotesDokumen7 halamanPrinciples of Inheritance and Variation-NotesVaibhav ChauhanBelum ada peringkat
- Andreaeales (Lantern Moss)Dokumen12 halamanAndreaeales (Lantern Moss)greeshma vasu100% (1)
- Understanding Plant Kingdom Through AlgaeDokumen19 halamanUnderstanding Plant Kingdom Through AlgaeDivyansha Sharma100% (1)
- Life Cycle of OedogoniumDokumen16 halamanLife Cycle of OedogoniumMuskan Sachdeva 0047Belum ada peringkat
- 5 Male Sterility 29-3-17Dokumen64 halaman5 Male Sterility 29-3-17Aizaz AliBelum ada peringkat
- Class 11 Chapter 3 BryophytesDokumen6 halamanClass 11 Chapter 3 BryophytesAshok KumarBelum ada peringkat
- 62e168e630d24 Plant Kingdom Mind MapDokumen3 halaman62e168e630d24 Plant Kingdom Mind MapDME MPonline100% (1)
- Introduction & Class of Pterido FinalDokumen19 halamanIntroduction & Class of Pterido FinalKritikaBelum ada peringkat
- Handouts-Lect. 29 - Introduction To Kingdom Plantae PDFDokumen6 halamanHandouts-Lect. 29 - Introduction To Kingdom Plantae PDFRizwan BalochBelum ada peringkat
- A Field Study Report On Education Trip From Sikkim To DarjeelingDokumen38 halamanA Field Study Report On Education Trip From Sikkim To DarjeelingVeekeshGuptaBelum ada peringkat
- EcosystemDokumen8 halamanEcosystemHarsha HinklesBelum ada peringkat
- Botany Marchantia Sem2Dokumen5 halamanBotany Marchantia Sem2Mega No01Belum ada peringkat
- Plant Tissue PDFDokumen63 halamanPlant Tissue PDFJitendra Kumar100% (1)
- General CharactersDokumen12 halamanGeneral CharactersKathirvel Gounder100% (1)
- Flower MorphologyDokumen129 halamanFlower MorphologySleeping Beauty100% (1)
- 044BIO11-03 Plant Kingdom (Study Notes)Dokumen10 halaman044BIO11-03 Plant Kingdom (Study Notes)Zen FredyBelum ada peringkat
- NCERT Filtrate 12 PAGESDokumen11 halamanNCERT Filtrate 12 PAGESKabir ThakurBelum ada peringkat
- Anthoceros ANTHOCEROS (Hornwort) : Morphology of ThallusDokumen3 halamanAnthoceros ANTHOCEROS (Hornwort) : Morphology of ThallusyeateshwarriorBelum ada peringkat
- 14 Respiration in Plants-NotesDokumen4 halaman14 Respiration in Plants-NotesAnanth DharanidharanBelum ada peringkat
- SpermatophytesDokumen26 halamanSpermatophytesNaomi Deirdre ReyesBelum ada peringkat
- Life Cycle of Pork TapewormDokumen13 halamanLife Cycle of Pork Tapeworms2s100% (1)
- Engler and Prantel System of ClassificationDokumen14 halamanEngler and Prantel System of ClassificationEasha tu RaziaBelum ada peringkat
- Principles of Inheritance and Variations: By: Dr. Anand ManiDokumen113 halamanPrinciples of Inheritance and Variations: By: Dr. Anand ManiIndu YadavBelum ada peringkat
- 12 Biology Notes Ch02 Sexual Reproduction in Flowering PlantsDokumen7 halaman12 Biology Notes Ch02 Sexual Reproduction in Flowering PlantsJatin kumar100% (1)
- Elements of GeneticsDokumen90 halamanElements of GeneticsArslan NazirBelum ada peringkat
- Gymnosperm (BOT-503) by Dr. Prabha DhondiyalDokumen59 halamanGymnosperm (BOT-503) by Dr. Prabha DhondiyalluckyBelum ada peringkat
- CH 7 - Structural Organisation in AnimalsDokumen8 halamanCH 7 - Structural Organisation in AnimalsLaxmiBelum ada peringkat
- CalobryalesDokumen7 halamanCalobryalesgreeshma vasuBelum ada peringkat
- Biology Study Material FinalDokumen157 halamanBiology Study Material FinalArush SharmaBelum ada peringkat
- The Living World Mind MapDokumen3 halamanThe Living World Mind MapDeepanyaBelum ada peringkat
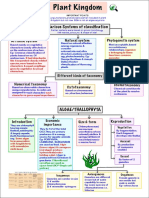
- Plant Kingdom Class 11 Notes NEET Biology (PDF)Dokumen17 halamanPlant Kingdom Class 11 Notes NEET Biology (PDF)shusa harshaBelum ada peringkat
- Excretion in Annelida: The Structure and Classification of NephridiaDokumen3 halamanExcretion in Annelida: The Structure and Classification of NephridiaNiroj Banik100% (1)
- Jim's Discussion Notes: How Plants Colonized LandDokumen21 halamanJim's Discussion Notes: How Plants Colonized LandSalik Sarosh TehamiBelum ada peringkat
- Class 11 Biology Chapter 4 NOTESDokumen38 halamanClass 11 Biology Chapter 4 NOTESYashica PradhanBelum ada peringkat
- RhyniopsidaDokumen3 halamanRhyniopsidax456456456xBelum ada peringkat
- BryophytaDokumen2 halamanBryophytaDiana SimbulBelum ada peringkat
- How Do Organisms Reproduce Notes (1) - EditedDokumen12 halamanHow Do Organisms Reproduce Notes (1) - EditedAALIYA ASHRAFBelum ada peringkat
- Lecture 21 The Stem (Primary Structure of Flowering Plant Body)Dokumen9 halamanLecture 21 The Stem (Primary Structure of Flowering Plant Body)maheen akramBelum ada peringkat
- Fungicides Passage Number 10 G/ML 1 Benomyl Continuous Aliette Roko Dhanuka RidomylDokumen9 halamanFungicides Passage Number 10 G/ML 1 Benomyl Continuous Aliette Roko Dhanuka RidomylvishcrimeBelum ada peringkat
- Sterilization Arabidopsis SeedDokumen1 halamanSterilization Arabidopsis SeedvishcrimeBelum ada peringkat
- Diversity of Seed Plants and Their Systematics by DR Nupur BhowmickDokumen26 halamanDiversity of Seed Plants and Their Systematics by DR Nupur Bhowmickdapolra55100% (2)
- Stomatal TypesDokumen5 halamanStomatal TypesvishcrimeBelum ada peringkat
- MSC Sub CommuDokumen64 halamanMSC Sub CommuvishcrimeBelum ada peringkat
- Ordinance No. 47Dokumen21 halamanOrdinance No. 47vishcrimeBelum ada peringkat
- Interimkey Set29may2016 PiiiDokumen124 halamanInterimkey Set29may2016 PiiivishcrimeBelum ada peringkat
- Statistical Tests in ExcelDokumen6 halamanStatistical Tests in Excelmak78Belum ada peringkat
- Murashige and Skoog Basal Salt MediumDokumen3 halamanMurashige and Skoog Basal Salt MediumvishcrimeBelum ada peringkat
- Contempt of Courts in IndiaDokumen31 halamanContempt of Courts in IndiaRattan WaraichBelum ada peringkat
- I. 7 PageDokumen7 halamanI. 7 PagevishcrimeBelum ada peringkat
- 313 Secondary GrowthDokumen21 halaman313 Secondary GrowthvishcrimeBelum ada peringkat
- Bioinfolet Seed OrnamentationDokumen4 halamanBioinfolet Seed OrnamentationvishcrimeBelum ada peringkat
- Seed Ornamentation Studies in Some Cleome SpeciesDokumen2 halamanSeed Ornamentation Studies in Some Cleome SpeciesvishcrimeBelum ada peringkat
- Effect of Powdery Mildew Infection On DPPH Radical Scavenging Activity and Ferric-Reducing Antioxidant Power of PlantsDokumen7 halamanEffect of Powdery Mildew Infection On DPPH Radical Scavenging Activity and Ferric-Reducing Antioxidant Power of PlantsvishcrimeBelum ada peringkat
- My BiodataDokumen8 halamanMy BiodatavishcrimeBelum ada peringkat
- Phytochemical ScreeningDokumen6 halamanPhytochemical ScreeningNkem OkonkwoBelum ada peringkat
- Phytochemical Analysis of WeedsDokumen4 halamanPhytochemical Analysis of WeedsvishcrimeBelum ada peringkat
- LogDokumen1 halamanLogvishcrimeBelum ada peringkat
- Phytochemical Screening of Commelina bengalensis and Cyanotis cristataDokumen3 halamanPhytochemical Screening of Commelina bengalensis and Cyanotis cristatavishcrimeBelum ada peringkat
- Books For PHDokumen1 halamanBooks For PHvishcrimeBelum ada peringkat
- Fiber Plants & DiscussionDokumen10 halamanFiber Plants & DiscussionvishcrimeBelum ada peringkat
- Insect PaperDokumen4 halamanInsect PapervishcrimeBelum ada peringkat
- Academic Audit 2010-2011Dokumen12 halamanAcademic Audit 2010-2011vishcrimeBelum ada peringkat
- KissDokumen2 halamanKissvishcrimeBelum ada peringkat
- San To SHDokumen2 halamanSan To SHvishcrimeBelum ada peringkat
- 111111Dokumen4 halaman111111vishcrimeBelum ada peringkat
- To Bio 1Dokumen18 halamanTo Bio 1Abdul RachmanBelum ada peringkat
- Life Cycle of A Bean Plant - Year 2 & 3Dokumen3 halamanLife Cycle of A Bean Plant - Year 2 & 3Regine BasilidesBelum ada peringkat
- Types of FruitsDokumen68 halamanTypes of FruitsUmang VermaBelum ada peringkat
- Vascular and Non-Vascular PlantsDokumen3 halamanVascular and Non-Vascular PlantsLeonardo PigaBelum ada peringkat
- George NakashimaDokumen6 halamanGeorge NakashimaCamiloJoséGonzalezObregon50% (2)
- Horticultural MCQ - S RepairedDokumen195 halamanHorticultural MCQ - S RepairedMik Khan0% (1)
- Plant StudyDokumen209 halamanPlant StudyScot RobertsBelum ada peringkat
- Soliva Anthemifolia PDFDokumen3 halamanSoliva Anthemifolia PDFSandeepBelum ada peringkat
- PLANTS REviewerDokumen3 halamanPLANTS REviewerJunie YeeBelum ada peringkat
- Module 6Dokumen5 halamanModule 6ZhajaBelum ada peringkat
- Botany Laboratory ManualDokumen22 halamanBotany Laboratory ManualKEVIN REY GUIRITANBelum ada peringkat
- Biology 12th Inner (Revised 2020-2021) - 1Dokumen344 halamanBiology 12th Inner (Revised 2020-2021) - 1Hitarth PatilBelum ada peringkat
- Anatomy of Nerium Leaf, Ovule and Anatomy of Anther 26.10.2019Dokumen2 halamanAnatomy of Nerium Leaf, Ovule and Anatomy of Anther 26.10.2019JAGGU BHAIBelum ada peringkat
- Monocot vs. Dicot: Roots: Fibrous vs. TaprootDokumen3 halamanMonocot vs. Dicot: Roots: Fibrous vs. TaprootExequiel ZoletaBelum ada peringkat
- Psdium Botanica 2012Dokumen12 halamanPsdium Botanica 2012Belén GuallichicoBelum ada peringkat
- AnthuriumDokumen17 halamanAnthuriumdhinglijay100% (2)
- Unit - IV Chapter-16. Photosynthesis: Important PointsDokumen16 halamanUnit - IV Chapter-16. Photosynthesis: Important PointsSantosh ChavanBelum ada peringkat
- Cell Structure and Function Multiple Choice Quiz Questions and AnswersDokumen32 halamanCell Structure and Function Multiple Choice Quiz Questions and AnswersAlyssa Kim Fernandez50% (4)
- Common diseases of important crops and their pathogensDokumen3 halamanCommon diseases of important crops and their pathogensAzriAzharBelum ada peringkat
- A New Species of Hampea Malvacea From El SalvadorDokumen4 halamanA New Species of Hampea Malvacea From El SalvadorG FuBelum ada peringkat
- BEGONIA Linnaeus, Sp. Pl. 2: 1056. 1753Dokumen56 halamanBEGONIA Linnaeus, Sp. Pl. 2: 1056. 1753Inayah Abdullah HasnyBelum ada peringkat
- Floral Biology, Objectives, Breeding Strategies and Achievements in Crop Improvement of RoseDokumen48 halamanFloral Biology, Objectives, Breeding Strategies and Achievements in Crop Improvement of RoseNithya Devi100% (1)
- Anatomy and Morphology of The Poaceae (Gramineae) - Grass FamilyDokumen43 halamanAnatomy and Morphology of The Poaceae (Gramineae) - Grass FamilyPeter B KihiyoBelum ada peringkat
- PlantsDokumen44 halamanPlantsEulalio NinaBelum ada peringkat
- PHT of Flower CropsDokumen55 halamanPHT of Flower CropsJagadish HortiBelum ada peringkat
- Some Beautiful Indian Climbers and ShrubsDokumen493 halamanSome Beautiful Indian Climbers and ShrubsMathew YoyakkyBelum ada peringkat
- Meristem Parenchyma Sclerenchyma 5: Figure 5: Cell Types and TissuesDokumen8 halamanMeristem Parenchyma Sclerenchyma 5: Figure 5: Cell Types and Tissuessandy waskitoBelum ada peringkat
- Sexual Reproduction in Flowering Plants (Note 2020) Part IDokumen11 halamanSexual Reproduction in Flowering Plants (Note 2020) Part IY S BAMMANAL y sBelum ada peringkat
- Effects of Different Light Intensities On Carbohydrate Accumulate and Distribution in Three Native Broad-Leaved Tree Species of TaiwanDokumen12 halamanEffects of Different Light Intensities On Carbohydrate Accumulate and Distribution in Three Native Broad-Leaved Tree Species of TaiwanJosephine ChiuBelum ada peringkat
- Agricultural Crops Production NC IIDokumen21 halamanAgricultural Crops Production NC IIsumaira abasBelum ada peringkat
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDari Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessPenilaian: 4 dari 5 bintang4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDari EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RacePenilaian: 4.5 dari 5 bintang4.5/5 (515)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDari EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeBelum ada peringkat
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDari EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindPenilaian: 4.5 dari 5 bintang4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDari EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondPenilaian: 4 dari 5 bintang4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyDari EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyPenilaian: 3.5 dari 5 bintang3.5/5 (31)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDari EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionPenilaian: 4 dari 5 bintang4/5 (811)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDari EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesPenilaian: 4.5 dari 5 bintang4.5/5 (397)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDari EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorBelum ada peringkat
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDari EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldPenilaian: 4.5 dari 5 bintang4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDari EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedPenilaian: 4 dari 5 bintang4/5 (11)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildDari EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildPenilaian: 4.5 dari 5 bintang4.5/5 (44)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineDari EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestinePenilaian: 4 dari 5 bintang4/5 (17)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishDari EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishPenilaian: 4 dari 5 bintang4/5 (30)
- Summary of Robert M. Sapolsky's DeterminedDari EverandSummary of Robert M. Sapolsky's DeterminedPenilaian: 5 dari 5 bintang5/5 (2)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDari EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsPenilaian: 4.5 dari 5 bintang4.5/5 (4)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesDari EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesPenilaian: 3.5 dari 5 bintang3.5/5 (56)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceDari EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceBelum ada peringkat
- Gathering Moss: A Natural and Cultural History of MossesDari EverandGathering Moss: A Natural and Cultural History of MossesPenilaian: 4.5 dari 5 bintang4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsDari EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsBelum ada peringkat
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationDari EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationPenilaian: 4 dari 5 bintang4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeDari EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifePenilaian: 4 dari 5 bintang4/5 (523)
- Superlative: The Biology of ExtremesDari EverandSuperlative: The Biology of ExtremesPenilaian: 4.5 dari 5 bintang4.5/5 (51)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDari EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsPenilaian: 4.5 dari 5 bintang4.5/5 (2083)
- Lymph & Longevity: The Untapped Secret to HealthDari EverandLymph & Longevity: The Untapped Secret to HealthPenilaian: 4.5 dari 5 bintang4.5/5 (13)