Anda mungkin juga menyukai
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- AP Psychology Module 66+67Dokumen5 halamanAP Psychology Module 66+67kyuuBelum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Netically Modified Crops Their Development, Uses, and Risks by G.H. LiangDokumen416 halamanNetically Modified Crops Their Development, Uses, and Risks by G.H. LiangbiokanagasundarBelum ada peringkat
- Management of DiabetesDokumen164 halamanManagement of Diabetesvivek_win95Belum ada peringkat
- Origin and Importance of GeneticsDokumen12 halamanOrigin and Importance of Geneticshuyenthanh1807100% (1)
- The Benefits of Cider VinegarDokumen15 halamanThe Benefits of Cider VinegarAldoSolsaBelum ada peringkat
- Medical Revision PaperDokumen106 halamanMedical Revision PaperDavin SugumarBelum ada peringkat
- Neonatal HyperglycemiaDokumen18 halamanNeonatal HyperglycemiayumicheongBelum ada peringkat
- Galvano-Spa-Bath and HealthDokumen2 halamanGalvano-Spa-Bath and HealthDeu na telha com Janaína BensdorpBelum ada peringkat
- 1 s2.0 S0828282X22001271 MainDokumen11 halaman1 s2.0 S0828282X22001271 MainLilianne Mbengani LaranjeiraBelum ada peringkat
- Review - Spinal Cord Injury - 2Dokumen1 halamanReview - Spinal Cord Injury - 2Sarah MendozaBelum ada peringkat
- Latihan Inggris SNMPTN 2012 Kode734Dokumen4 halamanLatihan Inggris SNMPTN 2012 Kode734Pamela FoxBelum ada peringkat
- Drug StudyDokumen14 halamanDrug StudyCj LowryBelum ada peringkat
- AUB - Physical Examination of UrineDokumen3 halamanAUB - Physical Examination of UrineJeanne RodiñoBelum ada peringkat
- Answers Part 2Dokumen64 halamanAnswers Part 2Rynjeff Lui-Pio100% (1)
- Physical Education and Health 4: Grade 12Dokumen39 halamanPhysical Education and Health 4: Grade 12Dio Emmanuel SadangBelum ada peringkat
- Oregon Trail DiaryDokumen4 halamanOregon Trail Diaryapi-262078158Belum ada peringkat
- IFDEA Dental Anatomy Educational Teaching ResourceDokumen38 halamanIFDEA Dental Anatomy Educational Teaching ResourcelienhuongahBelum ada peringkat
- Symptoms and Signs: Compartment SyndromeDokumen4 halamanSymptoms and Signs: Compartment SyndromeajeikoBelum ada peringkat
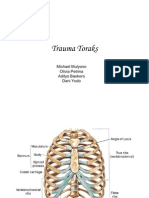
- Trauma ToraksDokumen30 halamanTrauma ToraksMichael KwanBelum ada peringkat
- BCAAs and Body Composition - Art de Vany On LineDokumen2 halamanBCAAs and Body Composition - Art de Vany On LineDellBelum ada peringkat
- Norsyahira Binti Abdullah - hs245 4cDokumen8 halamanNorsyahira Binti Abdullah - hs245 4cMOHD MU'IZZ BIN MOHD SHUKRIBelum ada peringkat
- Calamansi and Lemongrass As Alternative Mosquito Coil: Submitted By: Kristhel Nieves N. Zamora 10-AmethystDokumen6 halamanCalamansi and Lemongrass As Alternative Mosquito Coil: Submitted By: Kristhel Nieves N. Zamora 10-AmethystKristhel ZamoraBelum ada peringkat
- A Study On Knowledge and Practice of Mothers of Under-Five ChildrenDokumen6 halamanA Study On Knowledge and Practice of Mothers of Under-Five ChildrenRirin PurbaBelum ada peringkat
- Active Fluid De-Resuscitacion in Crtiticalli III Patients Whitj ShockDokumen8 halamanActive Fluid De-Resuscitacion in Crtiticalli III Patients Whitj ShockMartha Isabel BurgosBelum ada peringkat
- Jurnal ChikungunyaDokumen4 halamanJurnal ChikungunyaFitri Nadia SilvaniBelum ada peringkat
- Self-Injection Classes: Empowering Patients and Decreasing Nursing WorkloadDokumen5 halamanSelf-Injection Classes: Empowering Patients and Decreasing Nursing WorkloadIlvita MayasariBelum ada peringkat
- The Sovereign Remedy: Touch-Pieces and The King's Evil. (Pt. I) / Noel WoolfDokumen30 halamanThe Sovereign Remedy: Touch-Pieces and The King's Evil. (Pt. I) / Noel WoolfDigital Library Numis (DLN)Belum ada peringkat
- The Health Belief ModelDokumen13 halamanThe Health Belief ModelSteph DayaganonBelum ada peringkat
- Final Draft of Interview TranscriptDokumen7 halamanFinal Draft of Interview Transcriptapi-549369201Belum ada peringkat
- Genitourinary SystemDokumen27 halamanGenitourinary SystemDanica FrancoBelum ada peringkat