Anda mungkin juga menyukai
- Sistema NervosoDokumen1 halamanSistema NervosoPerisson Dantas100% (2)
- Rummy - RulesDokumen3 halamanRummy - RulesvijayBelum ada peringkat
- Student Number - Module Code - Question Number: Eg. 120456789 - BMS1234 - Q1Dokumen5 halamanStudent Number - Module Code - Question Number: Eg. 120456789 - BMS1234 - Q1Yusuf HasanBelum ada peringkat
- VaccinesDokumen39 halamanVaccinesritu9456Belum ada peringkat
- Triad Color TestDokumen37 halamanTriad Color TestC.O.M.A research -stopalienabduction-0% (1)
- DNA VaccinesDokumen14 halamanDNA VaccinesTrinity SSRBelum ada peringkat
- Journal Ppat 1000816Dokumen4 halamanJournal Ppat 1000816sanmusikBelum ada peringkat
- Editorial Overview Hildegund CJ Ertl and Antonio LanzavecchiaDokumen3 halamanEditorial Overview Hildegund CJ Ertl and Antonio Lanzavecchiahoangan118Belum ada peringkat
- Ru Ella 2013Dokumen25 halamanRu Ella 2013Ruben NatarîșBelum ada peringkat
- TMP 3941Dokumen16 halamanTMP 3941FrontiersBelum ada peringkat
- DNA Vaccination: Advanced Microbiology Course Prof: Rula DarwishDokumen30 halamanDNA Vaccination: Advanced Microbiology Course Prof: Rula Darwishmalak amerBelum ada peringkat
- Newgen VaccinesDokumen7 halamanNewgen Vaccinestranthuytrang213Belum ada peringkat
- Active Immunization Long DOCPDFDokumen31 halamanActive Immunization Long DOCPDFEnder IzaguirreBelum ada peringkat
- S1359644621002804Dokumen18 halamanS1359644621002804Natália FreitasBelum ada peringkat
- B and T Cell in ImmunologyDokumen6 halamanB and T Cell in ImmunologyTareque IslamBelum ada peringkat
- El Sulforafano FirstDokumen3 halamanEl Sulforafano FirstGiovanni Aleksey GuersonBelum ada peringkat
- Reviews: Modulating Immunity As A Therapy For Bacterial InfectionsDokumen12 halamanReviews: Modulating Immunity As A Therapy For Bacterial InfectionsZineb RAISBelum ada peringkat
- Vaccine Handbook Process Br5237en MKDokumen26 halamanVaccine Handbook Process Br5237en MKhadeer100% (2)
- Old and New Vaccine Approaches: Ruth Arnon, Tamar Ben-YedidiaDokumen10 halamanOld and New Vaccine Approaches: Ruth Arnon, Tamar Ben-YedidiahansmeetBelum ada peringkat
- Vaccine ImmunologyDokumen20 halamanVaccine ImmunologyAlejandro MotoliníaBelum ada peringkat
- VaccinesDokumen8 halamanVaccinesRaul MuresanBelum ada peringkat
- Vaccines For The 21st Century: ReviewDokumen13 halamanVaccines For The 21st Century: ReviewMuhafizBelum ada peringkat
- 1 s2.0 S0065266005540112 MainDokumen33 halaman1 s2.0 S0065266005540112 MainComodin PiterBelum ada peringkat
- Practical Applications of Immunology Written ReportDokumen9 halamanPractical Applications of Immunology Written ReportVincent Drystan AdronBelum ada peringkat
- Basic ImmunologyDokumen12 halamanBasic Immunologyjony_phurailatpamBelum ada peringkat
- Vaccines: Wim Jiskoot, Gideon F. A. Kersten, Enrico Mastrobattista, and Bram SlütterDokumen24 halamanVaccines: Wim Jiskoot, Gideon F. A. Kersten, Enrico Mastrobattista, and Bram SlütterSaritaJQM100% (1)
- Vaccination in Early LifeDokumen8 halamanVaccination in Early LifeLeonardo PalenciaBelum ada peringkat
- Activating Immunity To Fight A FoeDokumen3 halamanActivating Immunity To Fight A FoeGabriela ArvelaezBelum ada peringkat
- VaccinesDokumen8 halamanVaccinesArianaBelum ada peringkat
- Rakib L3 VaccinesDokumen23 halamanRakib L3 VaccinesSadia Afrin OshinBelum ada peringkat
- The Immune SystemDokumen4 halamanThe Immune SystemSseguya Lubega YaqubBelum ada peringkat
- TUBE MostCited 1Dokumen6 halamanTUBE MostCited 1jibin2008Belum ada peringkat
- Protection From HIV/AIDS: The Importance of Innate Immunity: Jay A. Levy, Iain Scott, and Carl MackewiczDokumen8 halamanProtection From HIV/AIDS: The Importance of Innate Immunity: Jay A. Levy, Iain Scott, and Carl MackewiczSofi Papel JachoBelum ada peringkat
- Vaccines - Definition, Principle, Types, Examples, Side EffectsDokumen26 halamanVaccines - Definition, Principle, Types, Examples, Side Effectssattyadev95Belum ada peringkat
- VACCINESDokumen8 halamanVACCINESJoyBelum ada peringkat
- Medicinal Chapter 5Dokumen4 halamanMedicinal Chapter 5WorthlessnessBelum ada peringkat
- Adaptações Na GravidezDokumen7 halamanAdaptações Na GravidezEstudanteBelum ada peringkat
- Artigo 3 - Com QuestionárioDokumen12 halamanArtigo 3 - Com QuestionárioLarissa SouzaBelum ada peringkat
- Edible Vaccines: Promises and Challenges: Vrinda M Kurup Jaya ThomasDokumen12 halamanEdible Vaccines: Promises and Challenges: Vrinda M Kurup Jaya ThomasBelaBelum ada peringkat
- 1 s2.0 S222116911530366X MainDokumen10 halaman1 s2.0 S222116911530366X MainNatália FreitasBelum ada peringkat
- AwerdtyuDokumen15 halamanAwerdtyuaycheewBelum ada peringkat
- Presentation (17) - 2Dokumen62 halamanPresentation (17) - 2ᏔᎬ'ᎯᎷ ᎯᏰᎠᎯᏞᏞᎻBelum ada peringkat
- Vaccination and Autoimmune Disease: What Is The Evidence?: ReviewDokumen8 halamanVaccination and Autoimmune Disease: What Is The Evidence?: Reviewtito7227Belum ada peringkat
- Novel Approaches For Vaccine Development: ReviewDokumen15 halamanNovel Approaches For Vaccine Development: ReviewAkula Sai PrasannaBelum ada peringkat
- Vaccine Development Strategies For Improving Immunization: The Role of Modern ImmunologyDokumen6 halamanVaccine Development Strategies For Improving Immunization: The Role of Modern ImmunologyVya PrabandariBelum ada peringkat
- What Are VaccinesDokumen9 halamanWhat Are VaccinesfaizanmuslimBelum ada peringkat
- Jansen 2019Dokumen9 halamanJansen 2019Roberto D. GarceteBelum ada peringkat
- Vaccin OlogyDokumen79 halamanVaccin Ologyhamzabinamjad987Belum ada peringkat
- 2015 FonsecaDokumen8 halaman2015 Fonsecacarlos tBelum ada peringkat
- Sysmex SEED Haematology InflammatoryDokumen7 halamanSysmex SEED Haematology InflammatoryTzeto Han CongBelum ada peringkat
- IMMUNIZATIONDokumen5 halamanIMMUNIZATIONbaker samuelBelum ada peringkat
- Tuberculosis: Lewis K. Schrager, Angelo Izzo, Kamalakannan VelmuruganDokumen5 halamanTuberculosis: Lewis K. Schrager, Angelo Izzo, Kamalakannan VelmurugancocorullBelum ada peringkat
- Immunization, Expanded Program On Immunization /universal Immunization Program ImmunizationDokumen21 halamanImmunization, Expanded Program On Immunization /universal Immunization Program ImmunizationHUSSAINA BANOBelum ada peringkat
- Nagy 2017Dokumen35 halamanNagy 2017Carlos ImasBelum ada peringkat
- Immunology of MycobacteriumDokumen37 halamanImmunology of MycobacteriumPhablo vinicius dos santos carneiroBelum ada peringkat
- WINSEM2021-22 BIY1020 TH VL2021220501009 Reference Material I 07-01-2022 3.immune Response Against A VaccineDokumen12 halamanWINSEM2021-22 BIY1020 TH VL2021220501009 Reference Material I 07-01-2022 3.immune Response Against A VaccineANENA RAJBelum ada peringkat
- 10 1056@NEJMra1911109 PDFDokumen11 halaman10 1056@NEJMra1911109 PDFMартин ДончевBelum ada peringkat
- 1 PBDokumen7 halaman1 PBEgo-free New-earthBelum ada peringkat
- C H N O: Cyclosporin ADokumen120 halamanC H N O: Cyclosporin AQfb Victor RobledoBelum ada peringkat
- 099 - VaccinesDokumen4 halaman099 - VaccinesArjan LallBelum ada peringkat
- VaccinesDokumen17 halamanVaccinesYOGANANTH mscasBelum ada peringkat
- Vaccination: How Millions of Lives Have Been Saved - Perhaps YoursDari EverandVaccination: How Millions of Lives Have Been Saved - Perhaps YoursBelum ada peringkat
- Management of Infections in the Immunocompromised HostDari EverandManagement of Infections in the Immunocompromised HostBrahm H. SegalBelum ada peringkat
- Erysipelothrix Rhusiopathiae: Molecular Medical Microbiology. DOI: © 2015 Elsevier Ltd. All Rights ReservedDokumen14 halamanErysipelothrix Rhusiopathiae: Molecular Medical Microbiology. DOI: © 2015 Elsevier Ltd. All Rights Reservedvignesh9489Belum ada peringkat
- Fungal Enzymes For Environmental Management: SciencedirectDokumen11 halamanFungal Enzymes For Environmental Management: SciencedirectsoultansimamoraBelum ada peringkat
- 4 261650026147611586 PDFDokumen19 halaman4 261650026147611586 PDFvignesh9489Belum ada peringkat
- Handbook of COVID-19 Prevention and Treatment (Compressed)Dokumen68 halamanHandbook of COVID-19 Prevention and Treatment (Compressed)Vanessa SánchezBelum ada peringkat
- Haemophilus InfluenzaeDokumen20 halamanHaemophilus Influenzaevignesh9489Belum ada peringkat
- GE AnswersDokumen1 halamanGE Answersvignesh9489Belum ada peringkat
- 4 261650026147611555 PDFDokumen9 halaman4 261650026147611555 PDFvignesh9489Belum ada peringkat
- Corynebacterium SPP, PDFDokumen7 halamanCorynebacterium SPP, PDFanggaririnBelum ada peringkat
- Haemophilus Ducreyi: Chancroid: EpidemiologyDokumen11 halamanHaemophilus Ducreyi: Chancroid: Epidemiologyvignesh9489Belum ada peringkat
- 4 261650026147611533 PDFDokumen15 halaman4 261650026147611533 PDFvignesh9489Belum ada peringkat
- 4 261650026147611555 PDFDokumen9 halaman4 261650026147611555 PDFvignesh9489Belum ada peringkat
- School RepresentativesDokumen4 halamanSchool Representativesvignesh9489Belum ada peringkat
- Enterotoxin-Producing Staphylococcus AureusDokumen17 halamanEnterotoxin-Producing Staphylococcus Aureusvignesh9489Belum ada peringkat
- Bacteriology of Periodontal DiseasesDokumen12 halamanBacteriology of Periodontal Diseasesvignesh9489Belum ada peringkat
- Lesson 15Dokumen13 halamanLesson 15Mystical MagicBelum ada peringkat
- Bacteriology of Periodontal DiseasesDokumen12 halamanBacteriology of Periodontal Diseasesvignesh9489Belum ada peringkat
- Corynebacterium SPP, PDFDokumen7 halamanCorynebacterium SPP, PDFanggaririnBelum ada peringkat
- Lec15 PDFDokumen47 halamanLec15 PDFvignesh9489Belum ada peringkat
- What Is DiabetesDokumen8 halamanWhat Is DiabetesJessica GintingBelum ada peringkat
- Candidemia PDFDokumen2 halamanCandidemia PDFRavi GulatiBelum ada peringkat
- E.coli Expression SystemsDokumen36 halamanE.coli Expression SystemstarannumBelum ada peringkat
- Picard Eau 2017Dokumen22 halamanPicard Eau 2017vignesh9489Belum ada peringkat
- 4 261650026147611533 PDFDokumen15 halaman4 261650026147611533 PDFvignesh9489Belum ada peringkat
- Gate 2018 Answer Key Architecture and PlanningDokumen3 halamanGate 2018 Answer Key Architecture and Planningvignesh9489Belum ada peringkat
- Entrance Test For M.Sc. (Biotechnology) I May 2L, 2oll IDokumen41 halamanEntrance Test For M.Sc. (Biotechnology) I May 2L, 2oll Ivignesh9489Belum ada peringkat
- Phosphate BufferDokumen1 halamanPhosphate BufferAdeel AltuhishBelum ada peringkat
- Bio - Technology Volume 10 Issue 12 1992 (Doi 10.1038 - nbt1292-1572) Fett-Neto, Arthur G. DiCosmo, Frank Reynolds, W. F. Sakata, K - Cell Culture of Taxus As A Source of The Antineoplastic Drug TaxDokumen4 halamanBio - Technology Volume 10 Issue 12 1992 (Doi 10.1038 - nbt1292-1572) Fett-Neto, Arthur G. DiCosmo, Frank Reynolds, W. F. Sakata, K - Cell Culture of Taxus As A Source of The Antineoplastic Drug Taxvignesh9489Belum ada peringkat
- Lab Manual For Biochemistry For PTU Students PDFDokumen24 halamanLab Manual For Biochemistry For PTU Students PDFvignesh9489Belum ada peringkat
- Qualitative Test of Lipids IIDokumen18 halamanQualitative Test of Lipids IIManohar Pattar75% (4)
- ANSA 019 SITXHRM001 Learner WorkbookDokumen42 halamanANSA 019 SITXHRM001 Learner WorkbookPalm ReaderBelum ada peringkat
- German Marine Agencies Inc Vs NLRC 142049 JanuaryDokumen14 halamanGerman Marine Agencies Inc Vs NLRC 142049 JanuaryJennyBelum ada peringkat
- Tr90 Flyer Eng (4 Pages)Dokumen4 halamanTr90 Flyer Eng (4 Pages)Cherry San Diego0% (1)
- HRM - CompleteDokumen12 halamanHRM - Completedhou_4Belum ada peringkat
- Health Economics: Which of The Following Is Not A Reason For Increased Health Spending?Dokumen8 halamanHealth Economics: Which of The Following Is Not A Reason For Increased Health Spending?Arjun Aryal100% (1)
- PHD Thesis On Coronary Artery DiseaseDokumen5 halamanPHD Thesis On Coronary Artery Diseasebdg9hkj6100% (1)
- BingggDokumen12 halamanBingggZulham YahyaBelum ada peringkat
- Journal Club: A Step Towards Evidence Based Practice: January 2011Dokumen6 halamanJournal Club: A Step Towards Evidence Based Practice: January 2011Jommy ChawalaBelum ada peringkat
- Country/Airport City Laboratory: AfghanistanDokumen22 halamanCountry/Airport City Laboratory: AfghanistanAliBelum ada peringkat
- Our Survival of SoulDokumen463 halamanOur Survival of SoulAnonymous Mq8EkiAyhM100% (2)
- Home Made ORSDokumen4 halamanHome Made ORSTrajan AureliusBelum ada peringkat
- Plazma Ionizarot CP 212 Eng ManualDokumen1 halamanPlazma Ionizarot CP 212 Eng ManualVladimirDulaBelum ada peringkat
- M265 Lec 13 - Epidemiology and Public HealthDokumen10 halamanM265 Lec 13 - Epidemiology and Public HealthSawsan Z. JwaiedBelum ada peringkat
- 07 RPM Chapter Feb 2021 V9 508 Clean FinalDokumen143 halaman07 RPM Chapter Feb 2021 V9 508 Clean FinalShubhamBelum ada peringkat
- Kacee-New ResumeDokumen4 halamanKacee-New Resumeapi-678307618Belum ada peringkat
- Mh25 Borderline Personality GuidelineDokumen182 halamanMh25 Borderline Personality GuidelineAbdul Hakim Abdul KadirBelum ada peringkat
- E Chlas Feb16Dokumen20 halamanE Chlas Feb16dkhatriBelum ada peringkat
- LCSW Additional CourseworkDokumen8 halamanLCSW Additional Courseworkiuhvgsvcf100% (2)
- NitrotolueneDokumen3 halamanNitrotolueneeriveraruizBelum ada peringkat
- Hardy Et Al-2018-Australian and New Zealand Journal of Public HealthDokumen5 halamanHardy Et Al-2018-Australian and New Zealand Journal of Public HealthAnggita RifkyBelum ada peringkat
- AyurvedaDokumen32 halamanAyurvedaRajeswari RanganathanBelum ada peringkat
- Ansdell G Elefant C. Pavlicevic M. Stige B. (2010) - Where Music Helps CommunityDokumen4 halamanAnsdell G Elefant C. Pavlicevic M. Stige B. (2010) - Where Music Helps CommunityNati de la BarraBelum ada peringkat
- MEM05052A Apply Safe Welding Practices - Learner GuideDokumen14 halamanMEM05052A Apply Safe Welding Practices - Learner Guidequestionbank.com.au100% (1)
- Claims FAQDokumen4 halamanClaims FAQSaiCharan GarimellaBelum ada peringkat
- Monkeypox: VirusDokumen18 halamanMonkeypox: Virusdiela dwi lestariBelum ada peringkat
- How Human Microbiome Talks To Health and DiseaseDokumen7 halamanHow Human Microbiome Talks To Health and DiseaseRafa SotoBelum ada peringkat
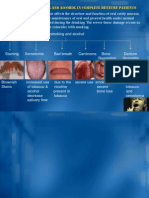
- Effects of Smoking and Alcohol On Complete Denture PatientsDokumen2 halamanEffects of Smoking and Alcohol On Complete Denture PatientsAshwin ThejaswiBelum ada peringkat
- Miranda Miracell NewDokumen2 halamanMiranda Miracell NewPablo FaldutiBelum ada peringkat