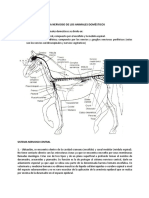
Anda mungkin juga menyukai
- Anatomia y Fisiología del Sistema Nervioso IIDari EverandAnatomia y Fisiología del Sistema Nervioso IIPenilaian: 4 dari 5 bintang4/5 (28)
- Sistema Nervioso de Los Pece1Dokumen3 halamanSistema Nervioso de Los Pece1Sonia Almendras QuispeBelum ada peringkat
- Manual Cto - EndocrinologíaDokumen145 halamanManual Cto - EndocrinologíaValeria Arauz Subiaga100% (5)
- Atlas Histología Vegetal y AnimalDokumen27 halamanAtlas Histología Vegetal y AnimalQuien S SabeBelum ada peringkat
- Sistema Nervioso Central para Estudio 1Dokumen5 halamanSistema Nervioso Central para Estudio 1Pilar Solas NuñezBelum ada peringkat
- El Sistema Nervioso en VertebradosDokumen2 halamanEl Sistema Nervioso en VertebradosAbdiel Haniel Coico LamaBelum ada peringkat
- Morfo2 (Recuperado Automáticamente)Dokumen20 halamanMorfo2 (Recuperado Automáticamente)Yarib Sarait VillarroelBelum ada peringkat
- Sistema NerviosoDokumen19 halamanSistema NerviosoErika GR100% (4)
- 12 Fisiología Sist. NerviosoDokumen10 halaman12 Fisiología Sist. NerviosoJasna ZovakBelum ada peringkat
- Informe de Ictiologia Semana #12Dokumen37 halamanInforme de Ictiologia Semana #12smaria2812Belum ada peringkat
- Sistemas Nerviosos en InvertebradosDokumen5 halamanSistemas Nerviosos en InvertebradosivanaBelum ada peringkat
- Sistema NerviosoDokumen9 halamanSistema NerviosoMarilyn AraujoBelum ada peringkat
- Taller de Fisiologia Sistema NerviosoDokumen20 halamanTaller de Fisiologia Sistema NerviosoLuz Mery Duran ZarateBelum ada peringkat
- Af - Unidad Nº10Dokumen8 halamanAf - Unidad Nº10jonathan caballeroBelum ada peringkat
- Presentación 3 Procesos de AprendizajeDokumen38 halamanPresentación 3 Procesos de AprendizajeJavier ValdezBelum ada peringkat
- Esquema Sistema Nervioso CentralDokumen7 halamanEsquema Sistema Nervioso CentralAngecrissBelum ada peringkat
- Protocolo 1 AnatomíaDokumen13 halamanProtocolo 1 AnatomíaJoanna HermosilloBelum ada peringkat
- Desarrollo Del Sistema NerviosoDokumen18 halamanDesarrollo Del Sistema NerviosoKaterine AvilaBelum ada peringkat
- MesensefaloDokumen10 halamanMesensefaloGaspar VelasquezBelum ada peringkat
- Anatomía Del Sistema Nervioso Central y Periférico Modificadas2Dokumen63 halamanAnatomía Del Sistema Nervioso Central y Periférico Modificadas2Georgina GalindoBelum ada peringkat
- TAREA 3.3 CUESTIONARIO SOBRE EL SNC Y EL SNP Denisse CabreraDokumen10 halamanTAREA 3.3 CUESTIONARIO SOBRE EL SNC Y EL SNP Denisse CabreraDenisse Cabrera ChalasBelum ada peringkat
- Sistema Nervioso CentralDokumen20 halamanSistema Nervioso CentralAlejandro TorrealbaBelum ada peringkat
- Par CranealDokumen179 halamanPar CranealRodrigo PonceBelum ada peringkat
- Qué Es El Encéfalo y Su FunciónesDokumen5 halamanQué Es El Encéfalo y Su FunciónesErika isamar Sánchez huamanBelum ada peringkat
- Presentacion Biologia GuionDokumen8 halamanPresentacion Biologia GuionEveBelum ada peringkat
- Practica #2 Reconocimiento Del Sistema Nervioso y Sus Principales EstructurasDokumen14 halamanPractica #2 Reconocimiento Del Sistema Nervioso y Sus Principales EstructurasCecilia Fernández JaramilloBelum ada peringkat
- G367 Tema 1. Organización Funcional Del Sistema NerviosoDokumen1 halamanG367 Tema 1. Organización Funcional Del Sistema NerviosoMaría GagoBelum ada peringkat
- Neurologia Ii: Bases Funcionales Del Sistema Nervioso Central Y PerifericoDokumen52 halamanNeurologia Ii: Bases Funcionales Del Sistema Nervioso Central Y PerifericoRocio AlbornozBelum ada peringkat
- 11 Sistema NerviosoDokumen5 halaman11 Sistema NerviosoAnder77Belum ada peringkat
- Trabajo de AnatomiaDokumen23 halamanTrabajo de AnatomiaLORENA VARGASBelum ada peringkat
- Introduccion Al CerebroDokumen6 halamanIntroduccion Al CerebroEDI8Belum ada peringkat
- Tipos de SN en AnimalesDokumen8 halamanTipos de SN en AnimalesJesus PitaBelum ada peringkat
- Sistema NerviosoDokumen10 halamanSistema NerviosoNikol Rodriguez Martinez 10Belum ada peringkat
- Resumen Y Mapas - SNC-SNP-y-Piel1-equipo 4Dokumen15 halamanResumen Y Mapas - SNC-SNP-y-Piel1-equipo 4Mauricio Caña AvalosBelum ada peringkat
- Sistema Nervioso CentralDokumen3 halamanSistema Nervioso CentralIrisBelum ada peringkat
- Sistema Nervioso y Niveles de OrganizacionDokumen11 halamanSistema Nervioso y Niveles de OrganizacionLizeth Ríos Caruajulca0% (1)
- Embriogenesis Del Sistema NerviosoDokumen19 halamanEmbriogenesis Del Sistema NerviosoYoselin Quispe vilcaBelum ada peringkat
- Medula OblongaDokumen9 halamanMedula OblongaLuis DavidBelum ada peringkat
- Sistema Nervioso 2 DiciembreDokumen7 halamanSistema Nervioso 2 DiciembreNathalie Fernanda ʚïɞBelum ada peringkat
- Glosario-Sistema NerviosoDokumen6 halamanGlosario-Sistema NerviosoSEGUNDO WUILMER MAJUAN NEYRABelum ada peringkat
- Sistema Nervios2Dokumen6 halamanSistema Nervios2Sofia GalvanBelum ada peringkat
- Anatomía Funcional Del Sistema NerviosoDokumen33 halamanAnatomía Funcional Del Sistema Nerviosoale192Belum ada peringkat
- Separata Sistema NerviosoDokumen17 halamanSeparata Sistema NerviosoVíctor Andres Mateo SanchezBelum ada peringkat
- Sistema NerviosoDokumen76 halamanSistema NerviosoJosué FloresBelum ada peringkat
- Bulbo Raquideo - CerebeloDokumen8 halamanBulbo Raquideo - CerebeloCINDY JESENIA MONTENEGRO MENABelum ada peringkat
- Cuestionario NeuroanatomíaDokumen20 halamanCuestionario NeuroanatomíaAlmaDieguezBelum ada peringkat
- 2 SEMA TEO Organizacion Del Sistema NerviosoDokumen17 halaman2 SEMA TEO Organizacion Del Sistema NerviosoEsmeralda De la cruz fabianBelum ada peringkat
- Fisio ContenidoDokumen444 halamanFisio ContenidojeronimoBelum ada peringkat
- Sistema NerviosoDokumen27 halamanSistema NerviosoEugenia VidelaBelum ada peringkat
- Desarrollo Filogenetico Del Sistema NerviosoDokumen13 halamanDesarrollo Filogenetico Del Sistema NerviosocpnvillasagricolasBelum ada peringkat
- Expo BiologiaDokumen5 halamanExpo Biologiastephanie erquinioBelum ada peringkat
- Sistema Nervioso CentralDokumen30 halamanSistema Nervioso CentralLeonardo Medina HornaBelum ada peringkat
- Tema 44: Órganos Y Funciones en Los Vertebrados 1. IntroducciónDokumen16 halamanTema 44: Órganos Y Funciones en Los Vertebrados 1. IntroducciónPaco BejaranoBelum ada peringkat
- El Sistema Nervioso Es Uno de Los Sistemas Más Importantes Del OrganismoDokumen21 halamanEl Sistema Nervioso Es Uno de Los Sistemas Más Importantes Del OrganismoMairelyn OvallesBelum ada peringkat
- Breve Resumen Sobre Neuroanatomia BasicaDokumen12 halamanBreve Resumen Sobre Neuroanatomia BasicaRosa GutierrezBelum ada peringkat
- ROMBENCÉFALODokumen15 halamanROMBENCÉFALOJazmín GarcíaBelum ada peringkat
- Sistema Nervioso AnatomíaDokumen82 halamanSistema Nervioso AnatomíaMariana RiekerBelum ada peringkat
- Sistema Nervioso AnimalDokumen3 halamanSistema Nervioso AnimalMel G100% (1)
- Resumen de Los Nervios Pares CranealesDokumen15 halamanResumen de Los Nervios Pares CranealesZunilda FleitasBelum ada peringkat
- Generalidades Del Sistema Nervioso CentralDokumen21 halamanGeneralidades Del Sistema Nervioso CentralLeise evelin Mamani huancaBelum ada peringkat
- Centros IntegradoresDokumen17 halamanCentros IntegradoresJorge DíazBelum ada peringkat
- 1 Molecula de AguaDokumen18 halaman1 Molecula de AguaMel LunaBelum ada peringkat
- Clase 1. 023Dokumen4 halamanClase 1. 023Mel LunaBelum ada peringkat
- El ACTO PUBLICO PARA LA DESIGNACION DE DOCENTES EN LA PROVINCIA DE BUENOS AIRESDokumen8 halamanEl ACTO PUBLICO PARA LA DESIGNACION DE DOCENTES EN LA PROVINCIA DE BUENOS AIRESMel LunaBelum ada peringkat
- Petróleo Y Sus DerivadosDokumen101 halamanPetróleo Y Sus DerivadosMel LunaBelum ada peringkat
- Tp3 Introduccion A La Quimica 5toDokumen4 halamanTp3 Introduccion A La Quimica 5toMel LunaBelum ada peringkat
- 6517 40618786 24 3 2020 PDFDokumen2 halaman6517 40618786 24 3 2020 PDFMel LunaBelum ada peringkat
- Guia 1 CENS188 Ciencias NaturalesDokumen5 halamanGuia 1 CENS188 Ciencias NaturalesMel LunaBelum ada peringkat
- Te 1606 PDFDokumen69 halamanTe 1606 PDFMel LunaBelum ada peringkat
- Cálculos de Entropía (GIE)Dokumen13 halamanCálculos de Entropía (GIE)Mel LunaBelum ada peringkat
- ChocolateDokumen18 halamanChocolateAsotibanaBelum ada peringkat
- Chocolate Belga PDFDokumen71 halamanChocolate Belga PDFAngel Rojas Swift100% (1)
- Estudio de La Germinación y La Conservación de Semillas de Cedro María (Calophyllum Brasiliense)Dokumen12 halamanEstudio de La Germinación y La Conservación de Semillas de Cedro María (Calophyllum Brasiliense)Enriz CosdaBelum ada peringkat
- CacaoDokumen302 halamanCacaoMel LunaBelum ada peringkat
- Cinetica ProblemasDokumen9 halamanCinetica ProblemasEduardoJoseBelum ada peringkat
- La Historia PDFDokumen2 halamanLa Historia PDFMel LunaBelum ada peringkat
- OgmDokumen69 halamanOgmMel LunaBelum ada peringkat
- Los Cultivos Transgénicos en Argentina y en El MundoDokumen17 halamanLos Cultivos Transgénicos en Argentina y en El MundoLuz Karina Pestana CaballeroBelum ada peringkat
- Material Didactico ReciclarDokumen4 halamanMaterial Didactico ReciclarMel LunaBelum ada peringkat
- Los Cultivos Transgénicos en Argentina y en El MundoDokumen17 halamanLos Cultivos Transgénicos en Argentina y en El MundoLuz Karina Pestana CaballeroBelum ada peringkat
- Justicia Nacional Electoral - Consulta Del Padrón Electoral PDFDokumen2 halamanJusticia Nacional Electoral - Consulta Del Padrón Electoral PDFMel LunaBelum ada peringkat
- Vermu THDokumen13 halamanVermu THMel LunaBelum ada peringkat
- Cart AmoDokumen1 halamanCart AmoMel LunaBelum ada peringkat
- Conograma Acto Público AdultosDokumen1 halamanConograma Acto Público AdultosAnibalPonceBelum ada peringkat
- La Alquimia 2013Dokumen2 halamanLa Alquimia 2013Mel LunaBelum ada peringkat
- La AlquimiaDokumen37 halamanLa AlquimiaKewin Joel Trujiillo BuitragoBelum ada peringkat
- Aldehidos y CetonasDokumen25 halamanAldehidos y Cetonasmarlu2424Belum ada peringkat
- La Quimica en La Vida CotidianaDokumen418 halamanLa Quimica en La Vida Cotidianaapi-373641880% (20)
- Las Mil y Una EmpanadasDokumen5 halamanLas Mil y Una EmpanadasMel LunaBelum ada peringkat
- Berazategui - La Contaminación Del Agua en El Río de La Plata Es - AlarmanteDokumen5 halamanBerazategui - La Contaminación Del Agua en El Río de La Plata Es - AlarmanteMel LunaBelum ada peringkat
- Tesis n0400 SorosDokumen38 halamanTesis n0400 SorosbrajhanBelum ada peringkat
- Apunte-La Cascada Del EstrésDokumen9 halamanApunte-La Cascada Del EstrésGaviotaBelum ada peringkat
- Evolucion de Los Problemas AmbientalesDokumen7 halamanEvolucion de Los Problemas AmbientalesWilliams Hernandez60% (5)
- Fisiología Humana-HormonasDokumen5 halamanFisiología Humana-HormonasAstrid Nicole Vega GonzálezBelum ada peringkat
- Carrizosa Hernández, Silvia. La Diferencia Sus Voces Ecos y Silencios PDFDokumen197 halamanCarrizosa Hernández, Silvia. La Diferencia Sus Voces Ecos y Silencios PDFSoledad Hernandez SolisBelum ada peringkat
- Trabajo HormonaDokumen51 halamanTrabajo HormonaNaira C Guerrero Del ToroBelum ada peringkat
- 1 - Fisiologia Del Sistema Endocrino Parte I-Mier26abr 2017-GvsDokumen24 halaman1 - Fisiologia Del Sistema Endocrino Parte I-Mier26abr 2017-GvsMariangeles quispe canaresBelum ada peringkat
- Clase 02 Eje Hipotalamo Hipofisis - Tum Hipof PDFDokumen113 halamanClase 02 Eje Hipotalamo Hipofisis - Tum Hipof PDFJair Alonso Paredes DezaBelum ada peringkat
- USMLE Step ENDOCRINODokumen33 halamanUSMLE Step ENDOCRINOleylanBelum ada peringkat
- Sistema EndocrinoDokumen5 halamanSistema EndocrinoAnderson OsorioBelum ada peringkat
- Tendencias CriminológicasDokumen64 halamanTendencias CriminológicasAlejandra OlmosBelum ada peringkat
- Resolución Guía de Repaso Sistema EndocrinoDokumen20 halamanResolución Guía de Repaso Sistema EndocrinoViviana Natalia GonzalezBelum ada peringkat
- Mini Ensayo PSU IIIDokumen11 halamanMini Ensayo PSU IIIConstanza CabañaBelum ada peringkat
- HipotálamoDokumen25 halamanHipotálamoTatiana ArgoteBelum ada peringkat
- 897 Puntos Par BiomagneticoDokumen53 halaman897 Puntos Par BiomagneticoChristiane Costa100% (1)
- Sistema DigestivoDokumen12 halamanSistema DigestivoAlfonsina AlbornozBelum ada peringkat
- Apunte Fisiologia II 2da ParteDokumen74 halamanApunte Fisiologia II 2da ParteJulieta De La RosaBelum ada peringkat
- Sesion 05 Sistema EndocrinoDokumen13 halamanSesion 05 Sistema EndocrinoAlex De La CruzBelum ada peringkat
- Biología - Tema 7 - La Relación en Los AnimalesDokumen9 halamanBiología - Tema 7 - La Relación en Los AnimalesMarta GarcíaBelum ada peringkat
- Sistema Endocrino de PerrosDokumen14 halamanSistema Endocrino de Perros-nevermind-Belum ada peringkat
- 2 Evaluación Sistema Nervioso y EndocrinoDokumen4 halaman2 Evaluación Sistema Nervioso y EndocrinojuanBelum ada peringkat
- Las Hormonas y Sus FuncionesDokumen13 halamanLas Hormonas y Sus FuncionesItzel C SantoyoBelum ada peringkat
- Neuroanatomia CerebroDokumen2 halamanNeuroanatomia CerebroSele SantanaBelum ada peringkat
- Como Funciona Nuestro Sistema Nervioso?: Ciencias 1Dokumen11 halamanComo Funciona Nuestro Sistema Nervioso?: Ciencias 1Osito Juan PandaBelum ada peringkat
- Principales Glándulas EndocrinasDokumen3 halamanPrincipales Glándulas EndocrinasNebur NochBelum ada peringkat
- El Sistema EndocrinoDokumen2 halamanEl Sistema EndocrinoSofia Jasmin Ccama MirandaBelum ada peringkat
- Carpeta de TrabajoDokumen133 halamanCarpeta de TrabajoErika Vela EstrellaBelum ada peringkat
- Clasificación de Las Glándulas y HormonasDokumen20 halamanClasificación de Las Glándulas y HormonasJosé DavidBelum ada peringkat
- Ciclo EstralDokumen7 halamanCiclo EstralLuis Fernelly Rodriguez DiazBelum ada peringkat
- Monografia Bases Biologicas Del Psiquimo HumanoDokumen45 halamanMonografia Bases Biologicas Del Psiquimo Humanogiovana churaBelum ada peringkat