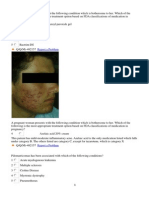
Anda mungkin juga menyukai
- General Dermatology MCQsDokumen197 halamanGeneral Dermatology MCQsDr.Tawheed95% (19)
- Case Study #5Dokumen2 halamanCase Study #5Jenny Jenders100% (1)
- Case Study 2.odtDokumen10 halamanCase Study 2.odtAlejandro Daniel Landa Morales100% (1)
- NCM 104 MusculoskeletalDokumen12 halamanNCM 104 MusculoskeletalCJ AngelesBelum ada peringkat
- Reflection About Patients With DementiaDokumen2 halamanReflection About Patients With Dementiaalexander abasBelum ada peringkat
- The Battle Against Covid-19 Filipino American Healthcare Workers on the Frontlines of the Pandemic ResponseDari EverandThe Battle Against Covid-19 Filipino American Healthcare Workers on the Frontlines of the Pandemic ResponseBelum ada peringkat
- Clinical Endocrinology of Dogs Cats PDFDokumen353 halamanClinical Endocrinology of Dogs Cats PDFsanjagruborovic100% (1)
- Biochem Mixed QuestionsDokumen220 halamanBiochem Mixed QuestionsAnonymous xk7cBdFBelum ada peringkat
- Dave Jay S. Manriquez RN. Acute CholecystitisDokumen11 halamanDave Jay S. Manriquez RN. Acute CholecystitisChilleMaeBelum ada peringkat
- Structured Health Teaching On Proper Nutrition of Common ConditionDokumen11 halamanStructured Health Teaching On Proper Nutrition of Common ConditionFry FryBelum ada peringkat
- SBAR ToolDokumen2 halamanSBAR Toolpragya_devkota0% (1)
- Evidence-Based Nursing: I. Clinical QuestionDokumen4 halamanEvidence-Based Nursing: I. Clinical QuestionRay Jorge MarmetoBelum ada peringkat
- Benign Prostatic HyperplasiaDokumen4 halamanBenign Prostatic Hyperplasiaエド パジャロンBelum ada peringkat
- Case 052: Biliary ColicDokumen4 halamanCase 052: Biliary ColicZauzaBelum ada peringkat
- Sexual Dysfunctions Self StudyDokumen6 halamanSexual Dysfunctions Self StudymelissagoldenBelum ada peringkat
- The Community Client: Assessment Diagnosis: As andDokumen43 halamanThe Community Client: Assessment Diagnosis: As andAbid HussainBelum ada peringkat
- Spinal Cord Injury Case Study (Physical Assessment)Dokumen3 halamanSpinal Cord Injury Case Study (Physical Assessment)TobiDaBelum ada peringkat
- Novilyn C. Pataray BSN - Ii: Assessment Diagnosis Pathophysiolog Y Planning Interevention Rationale EvaluationDokumen1 halamanNovilyn C. Pataray BSN - Ii: Assessment Diagnosis Pathophysiolog Y Planning Interevention Rationale EvaluationCharina AubreyBelum ada peringkat
- Prostate Cancer: Signs and SymptomsDokumen5 halamanProstate Cancer: Signs and SymptomsLicio LentimoBelum ada peringkat
- Compartment Syndrome: Return To TopDokumen3 halamanCompartment Syndrome: Return To TopSania Kamal BalweelBelum ada peringkat
- Bowel ObstructionDokumen6 halamanBowel ObstructionKryza Dale Bunado BaticanBelum ada peringkat
- Moira Mae B. Balite BSN 2A: Post Partum Care DefinitionDokumen5 halamanMoira Mae B. Balite BSN 2A: Post Partum Care DefinitionMoiraMaeBeridoBaliteBelum ada peringkat
- FNCPDokumen4 halamanFNCPKrista P. AguinaldoBelum ada peringkat
- Cervical Conization and The Risk of Preterm DeliveryDokumen11 halamanCervical Conization and The Risk of Preterm DeliveryGelo ConcepcionBelum ada peringkat
- Brief Description of The Disease/statistical IncidenceDokumen2 halamanBrief Description of The Disease/statistical IncidenceLeanne Princess Gamboa100% (1)
- Gouty Arthritis: Presented By: Petit Ivy Mae B. NacarioDokumen21 halamanGouty Arthritis: Presented By: Petit Ivy Mae B. NacarioMarivic DianoBelum ada peringkat
- The 8 D's of Stroke CareDokumen3 halamanThe 8 D's of Stroke Caremuhammad fahrizaBelum ada peringkat
- Prof Ad Day 1Dokumen136 halamanProf Ad Day 1Kareen ArnaizBelum ada peringkat
- Buerger's DiseaseDokumen4 halamanBuerger's DiseasesweetyjonasBelum ada peringkat
- Leukemias: Care SettingDokumen11 halamanLeukemias: Care SettingTinBelum ada peringkat
- Unit-Iii Gordon's Functional Health PatternsDokumen3 halamanUnit-Iii Gordon's Functional Health Patternsalphabennydelta4468Belum ada peringkat
- Malnutrition in Older AdultsDokumen44 halamanMalnutrition in Older Adultsjohnjoe subaynoBelum ada peringkat
- Discharge PlanDokumen4 halamanDischarge PlanKabang MoaBelum ada peringkat
- NCP DisturbedDokumen2 halamanNCP DisturbedTrixie Anne GamotinBelum ada peringkat
- 11 Digestion and Bowel Elimination 2019 (Nursing) Geriatric NursingDokumen44 halaman11 Digestion and Bowel Elimination 2019 (Nursing) Geriatric NursingDarla SaulerBelum ada peringkat
- Nusing CareplanDokumen3 halamanNusing Careplanardec_143Belum ada peringkat
- Cancer Case StudyDokumen33 halamanCancer Case StudyBorlongan PaulineBelum ada peringkat
- FNCP On Elevated Blood Pressure 2Dokumen4 halamanFNCP On Elevated Blood Pressure 2Aaron EspirituBelum ada peringkat
- NCM 107 1.2Dokumen2 halamanNCM 107 1.2Clint Mikael EulatrizBelum ada peringkat
- NCPDokumen10 halamanNCPCristina L. JaysonBelum ada peringkat
- Pathophysiology of Osteoporosis: AgingDokumen1 halamanPathophysiology of Osteoporosis: Agingnek100% (1)
- BSN 1 H Case Application Nursing Care PlanDokumen3 halamanBSN 1 H Case Application Nursing Care PlanAntonio EscotoBelum ada peringkat
- FractureDokumen71 halamanFractureRegime San Juan AtienzaBelum ada peringkat
- ABDOMINAL ObjectiveDokumen9 halamanABDOMINAL ObjectiveXing-Jin RomeroBelum ada peringkat
- Post Operative Care After AmputationDokumen11 halamanPost Operative Care After AmputationEka MahayundhariBelum ada peringkat
- Care PlanDokumen26 halamanCare PlanChevelle Valenciano-GaanBelum ada peringkat
- GI Diagnostic TestsDokumen7 halamanGI Diagnostic TestspatzieBelum ada peringkat
- Cardivascular Case Study 4Dokumen57 halamanCardivascular Case Study 4api-302598263Belum ada peringkat
- Epidemiology and Pathophysiology of Colonic Diverticular DiseaseDokumen8 halamanEpidemiology and Pathophysiology of Colonic Diverticular DiseaseAnonymous Hz5w55Belum ada peringkat
- Cholecystitis IntroductionDokumen4 halamanCholecystitis IntroductionJechelle Ann Pabustan Martin-BoniquitBelum ada peringkat
- ConceptMap AMLDokumen1 halamanConceptMap AMLnursing concept mapsBelum ada peringkat
- NCPDokumen4 halamanNCPJahmie RiveraBelum ada peringkat
- Effects of Aging On The Digestive SystemDokumen3 halamanEffects of Aging On The Digestive SystemShereen Manabilang100% (2)
- Case AnalysisDokumen12 halamanCase AnalysisFroilan TaracatacBelum ada peringkat
- In General, Amputation of Limbs Is TheDokumen11 halamanIn General, Amputation of Limbs Is TheCay Sevilla100% (1)
- A Simple Guide to Parathyroid Adenoma, Diagnosis, Treatment and Related ConditionsDari EverandA Simple Guide to Parathyroid Adenoma, Diagnosis, Treatment and Related ConditionsBelum ada peringkat
- Metabolic Alkalosis, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsDari EverandMetabolic Alkalosis, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsBelum ada peringkat
- Guidelines for Measuring Household and Individual Dietary DiversityDari EverandGuidelines for Measuring Household and Individual Dietary DiversityBelum ada peringkat
- Gastric Outlet Obstruction, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsDari EverandGastric Outlet Obstruction, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsBelum ada peringkat
- Avascular Necrosis, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsDari EverandAvascular Necrosis, A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsPenilaian: 4 dari 5 bintang4/5 (2)
- Ranula Cyst, (Salivary Cyst) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsDari EverandRanula Cyst, (Salivary Cyst) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsBelum ada peringkat
- CGJ 18 29Dokumen7 halamanCGJ 18 29ZenithaMeidaBelum ada peringkat
- CGJ 18 29 PDFDokumen6 halamanCGJ 18 29 PDFZenithaMeidaBelum ada peringkat
- S 019 LBLDokumen22 halamanS 019 LBLZenithaMeidaBelum ada peringkat
- Treatment of Steroid Sensitive Nephrotic Syndrome: A.S. AbeyagunawardenaDokumen8 halamanTreatment of Steroid Sensitive Nephrotic Syndrome: A.S. AbeyagunawardenaZenithaMeidaBelum ada peringkat
- Laporan Kasus Sindroma NefrotikDokumen13 halamanLaporan Kasus Sindroma NefrotikZenithaMeidaBelum ada peringkat
- Leukoaraiosis and StrokeDokumen5 halamanLeukoaraiosis and StrokeZenithaMeidaBelum ada peringkat
- What Are Heart Disease and Stroke?Dokumen2 halamanWhat Are Heart Disease and Stroke?ZenithaMeidaBelum ada peringkat
- Anatomy and Pathophysiology of Stroke: Eitan AurielDokumen8 halamanAnatomy and Pathophysiology of Stroke: Eitan AurielZenithaMeidaBelum ada peringkat
- PX NN CranialDokumen35 halamanPX NN CranialZenithaMeidaBelum ada peringkat
- Heart Failure in Pediatric Patients With Congenital Heart DiseaseDokumen18 halamanHeart Failure in Pediatric Patients With Congenital Heart DiseaseZenithaMeidaBelum ada peringkat
- Ist2008 PDFDokumen6 halamanIst2008 PDFZenithaMeidaBelum ada peringkat
- The Past TenseDokumen16 halamanThe Past TenseZenithaMeidaBelum ada peringkat
- Bab IDokumen4 halamanBab IZenithaMeidaBelum ada peringkat
- 1746 4358 4 13 PDFDokumen6 halaman1746 4358 4 13 PDFZenithaMeidaBelum ada peringkat
- Allopurinol and Bone Marrow AplasiaDokumen1 halamanAllopurinol and Bone Marrow AplasiaZenithaMeidaBelum ada peringkat
- Rabies Post Exposure Prophylaxis (RPEP) Algorithm: Call County Health DepartmentDokumen1 halamanRabies Post Exposure Prophylaxis (RPEP) Algorithm: Call County Health DepartmentZenithaMeidaBelum ada peringkat
- Intraoperative Parathyroid Hormone Assays - Uptodate 2022Dokumen18 halamanIntraoperative Parathyroid Hormone Assays - Uptodate 2022juanrangoneBelum ada peringkat
- K13-14 - Fisiologi Hormon Tiroid, Paratiroid, Dan AdrenalDokumen62 halamanK13-14 - Fisiologi Hormon Tiroid, Paratiroid, Dan AdrenalMuhammad Ikhram Habib DaulayBelum ada peringkat
- Disorders of Calcium and Phosphate MetabolismDokumen20 halamanDisorders of Calcium and Phosphate MetabolismAhmed Noureldin AhmedBelum ada peringkat
- Calcium, Phosphate and Vitamin D MetabolismDokumen25 halamanCalcium, Phosphate and Vitamin D MetabolismRendy LectusBelum ada peringkat
- Mrcog Sba's - 2Dokumen736 halamanMrcog Sba's - 2Histo Pathology86% (7)
- FULL-TEXT - NCLEX-RN Practice Quiz Test Bank 2 - NurseslabsDokumen88 halamanFULL-TEXT - NCLEX-RN Practice Quiz Test Bank 2 - NurseslabsRonaldo Matos PerezBelum ada peringkat
- Kab ReetDokumen254 halamanKab ReetAsia AlhkeemBelum ada peringkat
- Pathophysiology and Psychodynamics of Disease Causation, Homeostatic Mechanism, Life ProcessesDokumen13 halamanPathophysiology and Psychodynamics of Disease Causation, Homeostatic Mechanism, Life ProcessesMeenu Dogra67% (9)
- Endocrine SystemDokumen123 halamanEndocrine SystemZussette Corbita VingcoBelum ada peringkat
- David O. Norris, James A. Carr - Endocrine Disruption - Biological Bases For Health Effects in Wildlife and Humans-Oxford University Press, USA (2005) PDFDokumen492 halamanDavid O. Norris, James A. Carr - Endocrine Disruption - Biological Bases For Health Effects in Wildlife and Humans-Oxford University Press, USA (2005) PDFlashmiBelum ada peringkat
- Jose Maria College of MedicineDokumen35 halamanJose Maria College of MedicineRugene Naragas BeratoBelum ada peringkat
- Vitamina DDokumen18 halamanVitamina DFabrizio Antonilli MagroBelum ada peringkat
- Bone Metabolism: German VersionDokumen17 halamanBone Metabolism: German VersionDapot SianiparBelum ada peringkat
- Hypocalcemia CasesDokumen4 halamanHypocalcemia CasesRahmah Shah BahaiBelum ada peringkat
- Endocrine System Reviewer LecDokumen3 halamanEndocrine System Reviewer LecMadelleBelum ada peringkat
- 691 FTPDokumen101 halaman691 FTPAdel SherifBelum ada peringkat
- Vitamin DDokumen25 halamanVitamin DJawaherBelum ada peringkat
- Chapter 10 Hormones and Hormone Antagonists Complete PDF NotesDokumen12 halamanChapter 10 Hormones and Hormone Antagonists Complete PDF NotesJio InBelum ada peringkat
- Hipoparatiroidismo Postquirurgico. ATA 2018Dokumen12 halamanHipoparatiroidismo Postquirurgico. ATA 2018Nani HendrianiBelum ada peringkat
- Representative Laboratory Reference Values: Blood, Plasma, and SerumDokumen22 halamanRepresentative Laboratory Reference Values: Blood, Plasma, and SerumKubra ĖdrisBelum ada peringkat
- Pseudohypoparathyroidism-Literature Update 2018Dokumen8 halamanPseudohypoparathyroidism-Literature Update 2018radu nicolaeBelum ada peringkat
- Chapter 34 The Endocrine SystemDokumen39 halamanChapter 34 The Endocrine SystemCarl Agape Davis100% (3)
- Tablas Variabilidad Biológica Actualizadas 95% Confianza 2019Dokumen10 halamanTablas Variabilidad Biológica Actualizadas 95% Confianza 2019Sonrisa Posada RiañoBelum ada peringkat
- 100-Day Contract With Dairy CowDokumen22 halaman100-Day Contract With Dairy CowMarjorie SalgadoBelum ada peringkat
- Lecture Notes in Medical Technology - Lecture #6 - The PARATHYROID GLANDDokumen15 halamanLecture Notes in Medical Technology - Lecture #6 - The PARATHYROID GLANDKat JornadalBelum ada peringkat
- Summary - Blood Chemistry TestsDokumen38 halamanSummary - Blood Chemistry TestspayatingBelum ada peringkat
- 02 26 CCDokumen5 halaman02 26 CCJeanel SamonteBelum ada peringkat