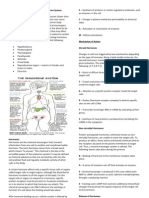
Anda mungkin juga menyukai
- endocrine فسيولوجياDokumen49 halamanendocrine فسيولوجياAUDEH AHMADBelum ada peringkat
- UntitledDokumen12 halamanUntitledDerrick kinyaBelum ada peringkat
- Endokrin MW DitransletDokumen7 halamanEndokrin MW DitransletSanta SaintBelum ada peringkat
- Endocrinology Notes: Veterinary PhysiologyDokumen16 halamanEndocrinology Notes: Veterinary PhysiologyBrian AllanBelum ada peringkat
- The Endocrine SystemDokumen37 halamanThe Endocrine SystemM Shafique SadiqeeBelum ada peringkat
- The Hypothalamus and Pituitary Gland: General Principles of Endocrine DiagnosisDokumen13 halamanThe Hypothalamus and Pituitary Gland: General Principles of Endocrine DiagnosisKiprono Keitany TimothyBelum ada peringkat
- Hormone NotesDokumen2 halamanHormone Notesapi-188978784100% (1)
- Endocrine ResonanceDokumen51 halamanEndocrine ResonanceEkta ManglaniBelum ada peringkat
- Module 1: The Endocrine System: Learning ObjectivesDokumen1 halamanModule 1: The Endocrine System: Learning ObjectivesZeina SharkasBelum ada peringkat
- Bio12 c10 10 2Dokumen10 halamanBio12 c10 10 2Eric McMullenBelum ada peringkat
- Clinton Hackney Anatomy & Physiology II BctcsDokumen65 halamanClinton Hackney Anatomy & Physiology II BctcsJack HolthouseBelum ada peringkat
- Endocrine Anatomy and PhysiologyDokumen10 halamanEndocrine Anatomy and PhysiologyJojo JustoBelum ada peringkat
- Clinical BiochemistryDokumen71 halamanClinical BiochemistryMuhamed ArsalanBelum ada peringkat
- Unit2-Hypothalamus and PituitaryDokumen80 halamanUnit2-Hypothalamus and Pituitarypranutan739Belum ada peringkat
- Phys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESDokumen7 halamanPhys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESsine summsun PradhanBelum ada peringkat
- ENDOCRINE SYSTEM - Anatomy and PhysiologyDokumen10 halamanENDOCRINE SYSTEM - Anatomy and PhysiologyJay Crishnan Morales CajandingBelum ada peringkat
- LECTURE 1 Endocrine Diseases Lecture 1 January 10-020Dokumen68 halamanLECTURE 1 Endocrine Diseases Lecture 1 January 10-020aleen qawareetBelum ada peringkat
- Endocrinology Apllied ScienceDokumen44 halamanEndocrinology Apllied ScienceAdam PrabowoBelum ada peringkat
- Endocrinedisorders Study 20session 201 1Dokumen39 halamanEndocrinedisorders Study 20session 201 1Yehuda Gelar PamungkasBelum ada peringkat
- Endocrine System: DR - Entissar Mansour 2O16-2017Dokumen162 halamanEndocrine System: DR - Entissar Mansour 2O16-2017Zainab Alshamary100% (1)
- Pituitary Hormones and Their Control by The HypothalamusDokumen20 halamanPituitary Hormones and Their Control by The HypothalamusMustafa KadhemBelum ada peringkat
- Unit IXDokumen10 halamanUnit IXPreeti ChouhanBelum ada peringkat
- Principles of Hormonal Control SystemDokumen25 halamanPrinciples of Hormonal Control SystemFAHEEM UD DINBelum ada peringkat
- Biochemistry of Hormones (Part 1)Dokumen31 halamanBiochemistry of Hormones (Part 1)arun231187100% (1)
- Endocrine System: Hypophysis Sella Turcica Pituitary HypophysialDokumen2 halamanEndocrine System: Hypophysis Sella Turcica Pituitary HypophysialMikaela MosqueraBelum ada peringkat
- EndocrineDokumen12 halamanEndocrinejamille consingBelum ada peringkat
- Bio ReportDokumen4 halamanBio ReportKate MabanagBelum ada peringkat
- Anatomy and Physiology-Endocrine SystemDokumen5 halamanAnatomy and Physiology-Endocrine SystemEixid Enna YeLikBelum ada peringkat
- Endocrine System Group 9Dokumen77 halamanEndocrine System Group 9Jei SanBelum ada peringkat
- Dmma College of Southern Philippines Anatomy and Physiology of Endocrine SystemDokumen16 halamanDmma College of Southern Philippines Anatomy and Physiology of Endocrine SystemCandy Penpillo SaldiviaBelum ada peringkat
- Care of Clients With Problem On EndocrineDokumen48 halamanCare of Clients With Problem On EndocrineAngel CauilanBelum ada peringkat
- Chapt 8 AnswersDokumen6 halamanChapt 8 AnswershulsercBelum ada peringkat
- Module 3Dokumen19 halamanModule 3VENTURA, ANNIE M.Belum ada peringkat
- Reproductive SystemDokumen114 halamanReproductive SystembookaccountBelum ada peringkat
- Hypothalamus: Done by Shahad Hamed 44003676Dokumen12 halamanHypothalamus: Done by Shahad Hamed 44003676shahad hamedBelum ada peringkat
- Endocrine GlandsDokumen36 halamanEndocrine GlandsnasyitahnorzlanBelum ada peringkat
- EN17 - Tumors of The Sellar Region - Acromegaly, Prolactinoma, Gonadotropinoma, Nonfunctioning Pituitary Adenoma, Craniopharyngioma)Dokumen24 halamanEN17 - Tumors of The Sellar Region - Acromegaly, Prolactinoma, Gonadotropinoma, Nonfunctioning Pituitary Adenoma, Craniopharyngioma)sbobine.imsBelum ada peringkat
- The Endocrine System 3 PDFDokumen47 halamanThe Endocrine System 3 PDFRifi OktasiaBelum ada peringkat
- Hypothalmo-Hypophysial Axis: by Dr. Luna PhukanDokumen35 halamanHypothalmo-Hypophysial Axis: by Dr. Luna PhukanAmmar QuasmiBelum ada peringkat
- Chemical Control and CoordinationDokumen19 halamanChemical Control and CoordinationShubhamBelum ada peringkat
- Lecture (Endocrine System)Dokumen15 halamanLecture (Endocrine System)GerofeBelum ada peringkat
- LMS Endocrine SystemDokumen23 halamanLMS Endocrine Systemhakdog hakdogBelum ada peringkat
- Pharmacology Chapter 34Dokumen5 halamanPharmacology Chapter 34languha NgatiBelum ada peringkat
- EndokrinDokumen70 halamanEndokrinNur Hadi PrayitnoBelum ada peringkat
- Human Anatomy & Physiology: Autonomic Nervous SystemDokumen8 halamanHuman Anatomy & Physiology: Autonomic Nervous SystemBeth EchavezBelum ada peringkat
- Unit 2 Part 1 Patho 2 Kmu IhsanDokumen33 halamanUnit 2 Part 1 Patho 2 Kmu IhsanRehan Khan RkBelum ada peringkat
- Chemical Control and IntegrationDokumen40 halamanChemical Control and IntegrationdiyBelum ada peringkat
- Drugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Dokumen467 halamanDrugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Marc Imhotep Cray, M.D.100% (1)
- Review Chapter 16 EndocrineDokumen11 halamanReview Chapter 16 EndocrineAkul MunjalBelum ada peringkat
- Unit 1 - Endocrine Guided NotesDokumen9 halamanUnit 1 - Endocrine Guided NotesJeanette IrambonaBelum ada peringkat
- Endocrine GlandDokumen4 halamanEndocrine Glandakash_zizouBelum ada peringkat
- Under The Guidance Of:: Dr. Sandeep Tandon Professor and Head of Dept. of PedodonticsDokumen79 halamanUnder The Guidance Of:: Dr. Sandeep Tandon Professor and Head of Dept. of PedodonticsMarivic DianoBelum ada peringkat
- Lecture 1, Introduction To EndocrinologyDokumen86 halamanLecture 1, Introduction To EndocrinologyAlexandru Ionut Popistaș100% (1)
- Endocrine PhysiologyDokumen32 halamanEndocrine PhysiologyAziem ShazwanBelum ada peringkat
- Hypothalamus - Anterior Pituitary and Their HormonesDokumen138 halamanHypothalamus - Anterior Pituitary and Their HormonesML Rodriguez100% (1)
- Topic 1 Endocrine SystemDokumen140 halamanTopic 1 Endocrine SystemmasdfgBelum ada peringkat
- Endocrine SystemDokumen69 halamanEndocrine SystemDivya JaiswalBelum ada peringkat
- Unit 13 Endocrine GlandDokumen73 halamanUnit 13 Endocrine GlandChandan ShahBelum ada peringkat
- Forelæsning 3Dokumen8 halamanForelæsning 3natashathelinBelum ada peringkat
- Endocrine Equilibrium: Navigating the Hormonal Seas: Navigating the Whispers of Hormones: A Delicate Dance of BalanceDari EverandEndocrine Equilibrium: Navigating the Hormonal Seas: Navigating the Whispers of Hormones: A Delicate Dance of BalanceBelum ada peringkat
- Hypothyroidism in PregnancyDokumen4 halamanHypothyroidism in PregnancyYesicaBelum ada peringkat
- Jurnal Hiperemesis GravidarumDokumen13 halamanJurnal Hiperemesis GravidarumfauziyahabidahBelum ada peringkat
- Adipose and Skeletal Muscle Thermogenesis: Studies From Large AnimalsDokumen17 halamanAdipose and Skeletal Muscle Thermogenesis: Studies From Large AnimalsBernard SoumaBelum ada peringkat
- Mcqs of Endocrine SystemDokumen25 halamanMcqs of Endocrine SystemkajuBelum ada peringkat
- 3 Use Thyroid Gland Use TTDokumen55 halaman3 Use Thyroid Gland Use TTMathy MtenjeBelum ada peringkat
- Pharmacology Block 4Dokumen83 halamanPharmacology Block 4Diwakesh C BBelum ada peringkat
- Sysemic Lupus Erythematosus Case StudyDokumen53 halamanSysemic Lupus Erythematosus Case StudyJennylen Torres100% (1)
- VPHY 143 Parathyroid To ThyroidDokumen3 halamanVPHY 143 Parathyroid To ThyroidRegulus Fidelis SevillaBelum ada peringkat
- Colloid Nodular GoiterDokumen37 halamanColloid Nodular GoiterLori GeorgeBelum ada peringkat
- SK Singh Lab Report 09042018Dokumen8 halamanSK Singh Lab Report 09042018Iasam Groups'sBelum ada peringkat
- Neonatology Pocket CambridgeDokumen601 halamanNeonatology Pocket CambridgeAnca Larisa92% (13)
- Pathogenesis of Hashimoto's Thyroiditis (Chronic Autoimmune Thyroiditis) - UpToDateDokumen22 halamanPathogenesis of Hashimoto's Thyroiditis (Chronic Autoimmune Thyroiditis) - UpToDaterazvanBelum ada peringkat
- Medicina 58 01115 v2Dokumen21 halamanMedicina 58 01115 v2silmanuryanzila2705Belum ada peringkat
- Ob Post TestDokumen11 halamanOb Post TestAlexBelum ada peringkat
- Labreportnew 6782Dokumen1 halamanLabreportnew 6782Shahabuddin AnsariBelum ada peringkat
- MicronutrientDokumen22 halamanMicronutrientSintayehu YeniewBelum ada peringkat
- 1 Pituitary and Thyroid PharmacologyDokumen64 halaman1 Pituitary and Thyroid PharmacologyZaina MasriBelum ada peringkat
- Hipo AsrticulDokumen6 halamanHipo AsrticulKarol MurciaBelum ada peringkat
- Endocrine Cases: Nahda College Program of Pharmacy Department of Clinical Pharmacy 2020-2021Dokumen29 halamanEndocrine Cases: Nahda College Program of Pharmacy Department of Clinical Pharmacy 2020-2021Basil DomiBelum ada peringkat
- Interview Jerry Tennant Healing Is VoltageDokumen19 halamanInterview Jerry Tennant Healing Is Voltagefoundryx2561100% (2)
- Applied Nutrition ManualDokumen205 halamanApplied Nutrition ManualLizzieBelum ada peringkat
- Coordination and Control: Animation 12.1: Neuron Source & Credit: Nichd - NihDokumen37 halamanCoordination and Control: Animation 12.1: Neuron Source & Credit: Nichd - Nihwardah nooriBelum ada peringkat
- What Is ManganeseDokumen2 halamanWhat Is ManganeseOm KarnBelum ada peringkat
- DR Lal Path Labs TestDokumen2 halamanDR Lal Path Labs TestPIVOT GRAVITYBelum ada peringkat
- Question For EndoDokumen30 halamanQuestion For EndoArti SharmaBelum ada peringkat
- Thyroid and Antityr DrugsDokumen23 halamanThyroid and Antityr Drugsmsmobile shBelum ada peringkat
- Emergencias Tiroideas 2019Dokumen17 halamanEmergencias Tiroideas 2019Marlon UlloaBelum ada peringkat
- Endocrine McQs 430Dokumen41 halamanEndocrine McQs 430MalikQamarAbbasThaheem83% (6)
- Euthrox CharacteristicsDokumen10 halamanEuthrox Characteristicspavaroti37Belum ada peringkat
- HLB, NME, M& M OSCE Notes - University of Manchester, WL GanDokumen45 halamanHLB, NME, M& M OSCE Notes - University of Manchester, WL GanWeh Loong Gan100% (2)