Anda mungkin juga menyukai
- Mreznica - 61Dokumen1 halamanMreznica - 61AkicaBelum ada peringkat
- Mreznica - 59Dokumen1 halamanMreznica - 59AkicaBelum ada peringkat
- Mreznica - 74Dokumen1 halamanMreznica - 74AkicaBelum ada peringkat
- Artigo Do Corey Sobre Adição de LiAlH4 em Alcoóis PropargílicosDokumen3 halamanArtigo Do Corey Sobre Adição de LiAlH4 em Alcoóis PropargílicosNatan FilippiBelum ada peringkat
- Tpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IDokumen1 halamanTpeak: I - Dark Adapted - 4"4 BG - 3.2 BG - 2.1 BG I I IAkicaBelum ada peringkat
- Mreznica - 64Dokumen1 halamanMreznica - 64AkicaBelum ada peringkat
- 4.3. Horizontal Cells: F I I FDokumen1 halaman4.3. Horizontal Cells: F I I FAkicaBelum ada peringkat
- Excitation: Function O'' Reaction C"Dokumen5 halamanExcitation: Function O'' Reaction C"anaklembah@ScribdBelum ada peringkat
- Divergence of The N-Loop Planar Graph in The Dual-Resonance Model (1971)Dokumen5 halamanDivergence of The N-Loop Planar Graph in The Dual-Resonance Model (1971)Mariangela GiulianeBelum ada peringkat
- First Page PDFDokumen1 halamanFirst Page PDFabdullah 3mar abou reashaBelum ada peringkat
- 102 2 21 Nanophysics Introduction Kwo EnglishDokumen96 halaman102 2 21 Nanophysics Introduction Kwo Englishquang_nh2007Belum ada peringkat
- Spin-Orbit Model. Evidence: Nuclear Configurations in The EmpiricalDokumen6 halamanSpin-Orbit Model. Evidence: Nuclear Configurations in The EmpiricalKaren lizeth fernanda Barbosa gomezBelum ada peringkat
- PhysRev 101 843Dokumen17 halamanPhysRev 101 843Maen SalmanBelum ada peringkat
- A Fresh Look at Longitudinal Standing Waves On A SpringDokumen4 halamanA Fresh Look at Longitudinal Standing Waves On A Spring林宸Belum ada peringkat
- L03 MRI Part 2Dokumen10 halamanL03 MRI Part 2lolpoll771Belum ada peringkat
- CHEM F111: General Chemistry: Semester I: AY 2017-18Dokumen17 halamanCHEM F111: General Chemistry: Semester I: AY 2017-18shrey shahBelum ada peringkat
- 6055d0ca2021 MidsemDokumen2 halaman6055d0ca2021 MidsemAnkit BishtBelum ada peringkat
- Ligero 2 Anghinolfi 1993Dokumen4 halamanLigero 2 Anghinolfi 1993Che ChoBelum ada peringkat
- APNFO4 YuDokumen2 halamanAPNFO4 YuLouis Chen100% (2)
- Kirmse 1984Dokumen3 halamanKirmse 1984Thiru98Belum ada peringkat
- Nonlinear Optical Properties of Organic Molecules and Crystals V1Dari EverandNonlinear Optical Properties of Organic Molecules and Crystals V1D.S. ChemlaBelum ada peringkat
- Nuclear Phy 2Dokumen31 halamanNuclear Phy 2Namitha RajeevBelum ada peringkat
- Edward Appleton - 1948 - Luxembourg Effect StudyDokumen33 halamanEdward Appleton - 1948 - Luxembourg Effect StudyMargot MA TAUPINBelum ada peringkat
- Membrane: Excitation Cardiac MuscleDokumen7 halamanMembrane: Excitation Cardiac MuscleNur Sa'idah044Belum ada peringkat
- Introduction To Nuclear PhysicsDokumen42 halamanIntroduction To Nuclear PhysicsRockiesBelum ada peringkat
- Mreznica - 80Dokumen1 halamanMreznica - 80AkicaBelum ada peringkat
- Kondalkar 2016Dokumen4 halamanKondalkar 2016lxvinh9467Belum ada peringkat
- documentDokumen8 halamandocumentstarseema312Belum ada peringkat
- B.sc. (H) Chemistry 1st Semester 2018Dokumen17 halamanB.sc. (H) Chemistry 1st Semester 2018Wajid PatelBelum ada peringkat
- Gomez Neuron95 CGMPDokumen12 halamanGomez Neuron95 CGMPJuan David Avila RinconBelum ada peringkat
- Radiative orbital electron capture in 193Pt analyzed to determine decay energyDokumen4 halamanRadiative orbital electron capture in 193Pt analyzed to determine decay energyRavi Sankar Babu BalabhadrapatruniBelum ada peringkat
- Berry - 1963 - The Reflectarray AntennaDokumen7 halamanBerry - 1963 - The Reflectarray Antennaanthony greavesBelum ada peringkat
- Radiation Loss 2Dokumen6 halamanRadiation Loss 2torikulBelum ada peringkat
- Blumenthal70 BHDokumen7 halamanBlumenthal70 BHStamatis StathBelum ada peringkat
- High-NOON States by Mixing Quantum and Classical LightDokumen4 halamanHigh-NOON States by Mixing Quantum and Classical LightItamar e Waneska sem sobrenomeBelum ada peringkat
- Nature 02199Dokumen4 halamanNature 02199Javi VRBelum ada peringkat
- 1974.12. Electromagnetic Fields Induced Inside Arbitrarily Shaped Biological BodyDokumen8 halaman1974.12. Electromagnetic Fields Induced Inside Arbitrarily Shaped Biological BodyEdy JjBelum ada peringkat
- Do Not: 5. Gain Control in PhotoreceptorsDokumen1 halamanDo Not: 5. Gain Control in PhotoreceptorsAkicaBelum ada peringkat
- Unit 5 Antenna and Wave PropagationDokumen18 halamanUnit 5 Antenna and Wave Propagationzathya darkBelum ada peringkat
- L04 MRI Part 3Dokumen21 halamanL04 MRI Part 3lolpoll771Belum ada peringkat
- 43 DecayDokumen9 halaman43 DecayMuhammad AhmadBelum ada peringkat
- The Transformation of Neuronal Activity Into Conscious Experience: The Syntergic Theory Jacobo GrinbergDokumen10 halamanThe Transformation of Neuronal Activity Into Conscious Experience: The Syntergic Theory Jacobo GrinbergsebastianBelum ada peringkat
- Simple Nanosecond Capacitive Voltage Divider: Additional Information On Rev. Sci. InstrumDokumen4 halamanSimple Nanosecond Capacitive Voltage Divider: Additional Information On Rev. Sci. InstrumdankorankoBelum ada peringkat
- Victor Novikov ITEP, Moscow, RussiaDokumen10 halamanVictor Novikov ITEP, Moscow, RussianouwezokanniewelwirBelum ada peringkat
- Locking of Hene Laser Modes Induced by Synchronous Intracavity ModulationDokumen3 halamanLocking of Hene Laser Modes Induced by Synchronous Intracavity ModulationMohamad paidarBelum ada peringkat
- Diffuse Reflectance Spectra and Optical Properties of Some Iron and Titanium Oxides and OxyhydroxidesDokumen8 halamanDiffuse Reflectance Spectra and Optical Properties of Some Iron and Titanium Oxides and Oxyhydroxidesicpsdt.cuetBelum ada peringkat
- 1907 04533 PDFDokumen6 halaman1907 04533 PDFKENET NARAY ROSALES FERREIRABelum ada peringkat
- Fabrication of High-Aspect-Ratio Silicon Nanostructures Using Near-Field Scanning Optical Lithography and Silicon Anisotropic Wet-Etching ProcessDokumen9 halamanFabrication of High-Aspect-Ratio Silicon Nanostructures Using Near-Field Scanning Optical Lithography and Silicon Anisotropic Wet-Etching ProcessDinesh LuniyaBelum ada peringkat
- Analysis: FrequenciesDokumen4 halamanAnalysis: FrequenciesOtman ChakkorBelum ada peringkat
- Cimoli No 1981Dokumen5 halamanCimoli No 1981Omar José Cotazo MosqueraBelum ada peringkat
- Cobalt Doped Nickel FerriteDokumen2 halamanCobalt Doped Nickel FerriteJeevan Job ThomasBelum ada peringkat
- LEDsDokumen61 halamanLEDsGeethakshayaBelum ada peringkat
- COL Circular Oblique LightingDokumen40 halamanCOL Circular Oblique LightingŁukasz OwsiankoBelum ada peringkat
- Nuclear Reactions and RadioactivityDokumen15 halamanNuclear Reactions and RadioactivityShannBelum ada peringkat
- Acustophoretic BasicsDokumen8 halamanAcustophoretic Basicsmechphysics21Belum ada peringkat
- Chemical Physics Lettres. 1990, 171, 209-216Dokumen8 halamanChemical Physics Lettres. 1990, 171, 209-216Jhony DeLas SalasBelum ada peringkat
- Ir Vis Uv Bb1998Dokumen8 halamanIr Vis Uv Bb1998Alejandro PuceiroBelum ada peringkat
- JJC 2008 H2 Chem - Atomic StructureDokumen16 halamanJJC 2008 H2 Chem - Atomic Structurebrianfletcher182100% (1)
- 1 C6cp07693aDokumen10 halaman1 C6cp07693aMd IsmayeelBelum ada peringkat
- Physics On Graphing Calculators: Articles You May Be Interested inDokumen3 halamanPhysics On Graphing Calculators: Articles You May Be Interested inbikash kumar naikBelum ada peringkat
- Mreznica - 80Dokumen1 halamanMreznica - 80AkicaBelum ada peringkat
- Mreznica - 79Dokumen1 halamanMreznica - 79AkicaBelum ada peringkat
- Mreznica - 78Dokumen1 halamanMreznica - 78AkicaBelum ada peringkat
- Mreznica - 71Dokumen1 halamanMreznica - 71AkicaBelum ada peringkat
- Et Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30Dokumen1 halamanEt Al. Macaca Fascicularis: 0 1 O0 200 3 O0 I I I 30AkicaBelum ada peringkat
- R. M. Shapley and C. Enroth-Cugell: Et AlDokumen1 halamanR. M. Shapley and C. Enroth-Cugell: Et AlAkicaBelum ada peringkat
- Visual Adaptation and Retinal Gain ControlsDokumen1 halamanVisual Adaptation and Retinal Gain ControlsAkicaBelum ada peringkat
- Z,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsDokumen1 halamanZ,, (Co) Y.L (Co) I, (O ) I, (Co) : Visual Adaptation and Retinal Gain ControlsAkicaBelum ada peringkat
- Mreznica - 73Dokumen1 halamanMreznica - 73AkicaBelum ada peringkat
- 6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oDokumen1 halaman6.1. Theories of The Retinal Gain Control: C O0 C - ' - ' - I C R oAkicaBelum ada peringkat
- Visual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)Dokumen1 halamanVisual Adaptation and Retinal Gain Controls Build Up of Particles ( 1 Y2 D Y3 LLLLL) L Y4) 1 Y5 ( Z,)AkicaBelum ada peringkat
- Mreznica - 64Dokumen1 halamanMreznica - 64AkicaBelum ada peringkat
- Mreznica - 66Dokumen1 halamanMreznica - 66AkicaBelum ada peringkat
- Retina - 52Dokumen1 halamanRetina - 52AkicaBelum ada peringkat
- Do Not: 5. Gain Control in PhotoreceptorsDokumen1 halamanDo Not: 5. Gain Control in PhotoreceptorsAkicaBelum ada peringkat
- Retina - 53 PDFDokumen1 halamanRetina - 53 PDFAkicaBelum ada peringkat
- Mreznica - 58Dokumen1 halamanMreznica - 58AkicaBelum ada peringkat
- 4.3. Horizontal Cells: F I I FDokumen1 halaman4.3. Horizontal Cells: F I I FAkicaBelum ada peringkat
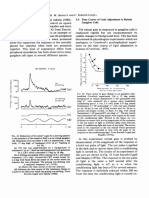
- Time Course of Gain Adjustment in Retinal Ganglion CellsDokumen1 halamanTime Course of Gain Adjustment in Retinal Ganglion CellsAkicaBelum ada peringkat
- Retina - 52Dokumen1 halamanRetina - 52AkicaBelum ada peringkat
- Visual Adaptation and Retinal Gain ControlsDokumen1 halamanVisual Adaptation and Retinal Gain ControlsAkicaBelum ada peringkat
- Retina - 52Dokumen1 halamanRetina - 52AkicaBelum ada peringkat
- Mreznica - 55Dokumen1 halamanMreznica - 55AkicaBelum ada peringkat
- Retina - 51Dokumen1 halamanRetina - 51AkicaBelum ada peringkat
- 3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsDokumen1 halaman3.6. Gain Control and Receptive Field Size Across The Population of Ganglion CellsAkicaBelum ada peringkat
- Topic 4 - Chemical Kinetics 4b - Half LifeDokumen20 halamanTopic 4 - Chemical Kinetics 4b - Half LifeJoshua LaBordeBelum ada peringkat
- 12 Translations PDFDokumen4 halaman12 Translations PDFTeo Lee Hong0% (1)
- Manage expenses and group contributions with Expense ManagerDokumen29 halamanManage expenses and group contributions with Expense ManagerPt Kamal SharmaBelum ada peringkat
- 4th Form Scheme of WorkDokumen6 halaman4th Form Scheme of WorkEustace DavorenBelum ada peringkat
- SSC JE Mechanical Study Material Strength of Materials PDFDokumen13 halamanSSC JE Mechanical Study Material Strength of Materials PDFSameer ShahBelum ada peringkat
- E8244 Ahu Catalogue 2014 PDFDokumen174 halamanE8244 Ahu Catalogue 2014 PDFmurugesanBelum ada peringkat
- Web Intelligence XI 3.0 Parameter GuideDokumen10 halamanWeb Intelligence XI 3.0 Parameter GuideJaime Andrés Triviño Sánchez0% (1)
- Continuous DeploymentDokumen2 halamanContinuous DeploymentVenkat Reddy0% (1)
- C - Brim - 1909 80 - QDokumen40 halamanC - Brim - 1909 80 - Qmanohar rathodBelum ada peringkat
- Lab 8 - LP Modeling and Simplex MethodDokumen8 halamanLab 8 - LP Modeling and Simplex MethodHemil ShahBelum ada peringkat
- Staad FoundationDokumen25 halamanStaad FoundationAnonymous nwByj9L100% (2)
- Parker 1991Dokumen17 halamanParker 1991Rodrigo GarcíaBelum ada peringkat
- Celda Carga - KIS-2BA - 5KNDokumen4 halamanCelda Carga - KIS-2BA - 5KNJosé TimanáBelum ada peringkat
- Lecture Antenna MiniaturizationDokumen34 halamanLecture Antenna MiniaturizationJuhi GargBelum ada peringkat
- Open Problems in The Mathematics of Data ScienceDokumen152 halamanOpen Problems in The Mathematics of Data Sciencecan dagidirBelum ada peringkat
- Unit 2Dokumen76 halamanUnit 2Mithila100% (1)
- Petroleum GeomechanicsDokumen35 halamanPetroleum GeomechanicsAnonymous y6UMzakPW100% (1)
- Sampling, Testing, and Analysis of Asphalt Roll Roofing, Cap Sheets, and Shingles Used in Roofing and WaterproofingDokumen13 halamanSampling, Testing, and Analysis of Asphalt Roll Roofing, Cap Sheets, and Shingles Used in Roofing and WaterproofingLuigi HernándezBelum ada peringkat
- Breathing AND Exchange of Gases MCQs PDFDokumen78 halamanBreathing AND Exchange of Gases MCQs PDFJatin SinglaBelum ada peringkat
- Microelectronics: Circuit Analysis and Design, 4 Edition by D. A. Neamen Problem SolutionsDokumen6 halamanMicroelectronics: Circuit Analysis and Design, 4 Edition by D. A. Neamen Problem SolutionsJano Jesus AlexBelum ada peringkat
- Complex Numbers: Problem-SolvingDokumen2 halamanComplex Numbers: Problem-SolvingK TariBelum ada peringkat
- Cs8080 - Irt - Notes AllDokumen281 halamanCs8080 - Irt - Notes Allmukeshmsd2Belum ada peringkat
- Subsurface Remote Sensing of Kelp ForestsDokumen20 halamanSubsurface Remote Sensing of Kelp Forestsjpeterson1Belum ada peringkat
- Basic Symbols and Fundamental Elements of Technical Drawing 1Dokumen26 halamanBasic Symbols and Fundamental Elements of Technical Drawing 1seanmatthewferrerBelum ada peringkat
- SKM and OKM User ManualDokumen36 halamanSKM and OKM User ManualOutlawRBelum ada peringkat
- 14GMK 6250 TelescopeDokumen13 halaman14GMK 6250 TelescopeВиталий РогожинскийBelum ada peringkat
- 10th Geometrical ConstructionDokumen2 halaman10th Geometrical Constructionapi-242227794100% (1)
- Seamless Steel Tubes and Pipes For BoilersDokumen20 halamanSeamless Steel Tubes and Pipes For BoilersABChungBABelum ada peringkat
- Standard Normal Distribution Table PDFDokumen1 halamanStandard Normal Distribution Table PDFWong Yan LiBelum ada peringkat
- Shipboard: Micrpoclimate' Coolin SysemsDokumen47 halamanShipboard: Micrpoclimate' Coolin Sysemsjakalae5263Belum ada peringkat